
Виды сорняков: название, описание, фото
Один из наиболее важных пунктов по уходу за огородными грядками, садом и газоном – профилактика и удаление сорняков. Способы борьбы зависят от вида сорного растения, его биологических особенностей и места произрастания. Рассмотрим основные типы сорняков и опишем методы контроля над их распространением.
Содержание
- Вред и польза от сорных растений
- Классификация сорняков
- Cорные растения на огороде: названия, описания, фото
- Газонные сорняки: названия, описания, фото
- Полезные сорняки на участке
- Профилактика появления сорняков
Вред и польза от сорных растений
Сорняками принято называть растения, которые «поселились» на участке помимо возделываемых культур. Борьба с ними отнимает у дачников очень много времени и сил. На сегодняшний день известно более 2000 сорных растений, в том числе ядовитых и вредных для животных – около 100 наименований.
Как бы мы ни старались, но избавиться один раз и навсегда от сорных растений не получиться – они попадают на участок через самые разные источники:
- некоторые семена сорняков «сидят» в почве и ожидают благоприятных условий для прорастания;
- органические удобрения – если компост не прошел должную обработку;
- некачественный посевной материал;
- семена заносятся ветром, животными, людьми (на подошве обуви).
Сорные растения причиняют ощутимый вред земледелию и ландшафтному дизайну:
- заглушают культурные растения и снижают их урожайность;
- выделяют в почву вредные вещества;
- поглощают большое количество питательных веществ и воды;
- создают тень;
- могут стать причиной отравления домашних животных;
- являются очагом для развития болезней и вредителей с/х культур.
Но, не все так однозначно. Агротехники отмечают, что некоторые виды сорных растений приносят и пользу.
Классификация сорняков
Все сорные растения классифицируют по трем основным биологическим признакам:
- продолжительность жизни;
- способ размножения;
- способ питания.
Зависимо от продолжительности жизни сорняки подразделяют на малолетние и многолетние виды.
Малолетние «бурьяны» размножаются семенами. Эта группа включает в себя:
- эфемеры – вегетационный период меньше одного сезона;
- яровые – вегетационный период такой же, как и у однолетних огородных растений; чаще остальных сорняков засоряют культурные посевы;
- озимые однолетние – всходят в начале осени; засоряют посадки многолетних трав и посевы пшеницы;
- двулетние – полный цикл развития включает два вегетационных периода.


Многолетние сорняки могут расти на одном месте до 4-х лет. После того, как семена созревают, наземные органы растения отмирают, а корневая система продолжает развиваться. Ежегодно от подземной части отрастают новые стебли. Многолетники размножаются вегетативно или семенами.
По способу питания различают такие виды сорняков:
- Непаразитные сорняки имеют самостоятельный тип питания и их развитие не зависит от других растений. Наиболее многочисленная группа.
- Полупаразитные сорняки имеют способность к фотосинтезу, но в тоже время, частично питаются за счет других культур – присасываются к наземным органам или корням растений. Типичные представители: ремнецветник европейский, омела белая, марьянник полевой, мытник болотный, зубчатка поздняя.
- Паразитные сорняки – растение не имеет зеленых листьев, корней, не способно к фотосинтезу и питается исключительно за счет другого растения. Сорняки-паразиты крепятся к корням или стеблям культурных растений. К этой группе относятся: повилика клеверная, повилика льняная, заразиха подсолнечниковая. Способ размножения – семенами.
 К этой группе относятся: повилика клеверная, повилика льняная, заразиха подсолнечниковая. Способ размножения – семенами.
К этой группе относятся: повилика клеверная, повилика льняная, заразиха подсолнечниковая. Способ размножения – семенами. Сорные растения на огороде: названия, описания, фото
Пырей ползучий обитает на полях, огородах, в садах, в поймах рек, вдоль дорог. Имеет глубокую корневую систему и быстро распространяется по участку. Стебель сорняка – прямостоячий, листья – плоские, длинные с шероховатой поверхностью. Пырей очень устойчив к неблагоприятным условиям, размножается корневищами практически на любом типе почве, многолетнее растение семейства злаковых. От пырея надо избавляться, особенно если планируется посадка картофеля.
Если не бороться с пыреем, газон может полностью зарости сорняками: фото
Вьюнок полевой (березка) – многолетний сорняк, обволакивающий стебли растений. Одно растение способно запутать до 2-х квадратных метров площади посевов. Больше всего от березки страдают ягодные кустарники. Длина стебля – до 180 см, листья – стреловидные, плод – двухгнездная коробочка. Разветвленные корни полевого вьюнка уходят в землю на глубину до пяти метров, поэтому полностью избавиться от растения очень сложно – корни придется выкапывать.
Больше всего от березки страдают ягодные кустарники. Длина стебля – до 180 см, листья – стреловидные, плод – двухгнездная коробочка. Разветвленные корни полевого вьюнка уходят в землю на глубину до пяти метров, поэтому полностью избавиться от растения очень сложно – корни придется выкапывать.
Портулак огородный – однолетнее растение с красноватым толстым стеблем и мясистыми листьями. Длина стебля – около 60 см, побеги одного растения могут покрыть значительную площадь огорода. Портулак может использоваться в лекарственных целях и в кулинарии.
Мокрица (звездчатка) – зимующий однолетник, эфемер. Всходы сорняка появляются ранней весной, а когда наступает время для прорастания культурных растений – мокрица разрастается в сплошной зеленый ковер. От этого сорняка сильно страдают посевы моркови.
В старину по мокрице предсказывали погоду на ближайшее время. Считалось, что если после восхода солнца цветки звездчатки не поднимались и не раскрывались, то днем следует ждать осадков
Щирица запрокинутая – яровый ранний однолетник, отличающийся очень высокой плодовитостью.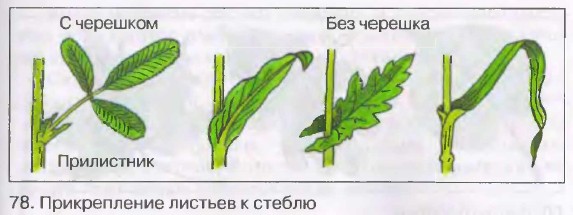 Семеня сорняка не боятся механических воздействий и сохраняют способность к прорастанию на протяжении 5-40 лет. Растение имеет высокий опушенный стебель (до 150 см), яйцевидно-ромбические листья, цветки собраны в густое метельчатое соцветие. Семена могут прорасти с глубины не больше 3 см.
Семеня сорняка не боятся механических воздействий и сохраняют способность к прорастанию на протяжении 5-40 лет. Растение имеет высокий опушенный стебель (до 150 см), яйцевидно-ромбические листья, цветки собраны в густое метельчатое соцветие. Семена могут прорасти с глубины не больше 3 см.
Ежовник (куриное просо) – засоряет овощные культуры (подсолнечник, морковь, свеклу) в начале их роста. В дождливые года способен полностью заглушить изреженные молодые посевы. Стебель достигает 120 см в высоту, листья – широколинейные, заостренные по краям, соцветие – метелка с колючими одноцветковыми колосками, наполненная семенами, которые похожи на просо.
Подмаренник цепкий – ранний яровой однолетник. Отличительная особенность – ствол и листья сорняка буквально цепляются за одежду. Подмаренник цепкий появляется на плодородных, богатых известняком почвах. Коневая система сорняка – стержневая, стебель – четырехгранный высотою до 1 м, на ребрах стебля есть маленькие, загнутые вниз шипы. Цветки собраны в густые метелки, период цветения – лето.
Цветки собраны в густые метелки, период цветения – лето.
Пастушья сумка
– зимующий однолетник, цветущий с весны до поздней осени. Жизнеспособность семян сохраняется почти 35 лет. Высота стебля около 20-40 см, листья перисто-рассеченной формы, корень – стержневой. Белые мелкие цветки собраны на самой верхушке стебля, цветение сорняка продолжается все лето. За это время растение дает 2-4 поколения (семена опадают и сразу же прорастают).Ярутка полевая – однолетнее растение с характерными округлыми плодами с вырезом на верхушке. Высота стебля – не более 40 см, листья собраны в розетку и находятся у земли. За период цветения ярутка дает до 50 тысяч семян. Максимальная глубина, с которой семена могут прорости – 5 см.
Осот розовый, более известный, как бодяк полевой – распространенное многолетнее растение сорняк, засоряющее любые посевы. Высота осота может достигать 1,5 м, поверхность стебля колючая. Форма листа ланцетная, по краям имеются колючки.
Основной источник засорения огорода бодяком – вертикальный корень. При борьбе с осотом необходимо уничтожать корневую систему, расположенную на глубине до 60-70 см
Газонные сорняки: названия, описания, фото
Мятлик – однолетний сорняк. В начале роста растение незаметно, но со временем сорняк зацветает и выделяется некрасивыми пятнами на газонном ковре. Мятлик хорошо себя чувствует на уплотненных почвах в низинах. Если своевременно вывести сорняк, то он долго не будет появляться на газоне.
Не избежать появления на газоне одуванчиков, семена которых разносятся ветром. Лучше всего одуванчики «приживаются» на молодых, редко засаженных газонах. Сорное растение имеет мясистые корни, в которых содержится большой запас питательных веществ. Это надо учитывать при борьбе с сорняком – для полного уничтожения понадобится не одна обработка гербицидами избирательного действия.
Лютик ползучий – многолетний сорняк со стелющимся по земле стеблем. Длина растения – около 1 м, листья имеют тройчатую форму. Лютик размножается семенами и вегетативно. Благоприятно развивается во влажных и недостаточно освещенных местах.
Мох, как и лютик, появляется на сырых почвах в низинах. Для борьбы с ним газон надо регулярно аэрировать и делать дренажные канавы. Высота мха – не более 50 см. Если не бороться с этим сорняком, то он способен полностью вытеснить газонные злаки и привести к заболачиванию почвы.
Появление мха может свидетельствовать о «бедном» составе почвы, нехватке питательных элементов и чрезмерном уровне кислотности грунта
Подорожник
Вероника нитчатая селится на увлажненной почве, богатой на полезные элементы. Высота растения – не более 12 см, стебель тонкий, цветы нежно лилового цвета. Сорняк размножается побегами.
Клевер – наиболее проблемный сорняк, доставляющий массу хлопот владельцам газонов. Многолетнее растение высотою 15-50 см, корневая система – стержневая, листья яйцевидной формы. Появление клевера может сигнализировать о нехватке в почве азота.
Активный рост клевера могут спровоцировать калийные и фосфорные удобрения, внесенные весной
Марь белая — морозоустойчивый сорняк на участке, растущий с ранней весны до глубокой осени. Высота растения может достигать 1,5 м. Максимальная глубина, с которой семена могут прорости – 10 см. Растение надо удалять до цветения, так как один сорняк может дать около 500 тысяч семян с различными сроками всходов.
Кислица обыкновенная (заячья капуста)– многолетнее растение с мощной корневой системой. Сорняк растет группами и с легкостью «забивает» газонную траву. Кислица очень устойчива к многим химическим препаратам, поэтому наиболее действенный способ – вырывание сорняка с корнем.
Сорняк растет группами и с легкостью «забивает» газонную траву. Кислица очень устойчива к многим химическим препаратам, поэтому наиболее действенный способ – вырывание сорняка с корнем.
Представленные фото и названия сорняков помогут вовремя распознать и искоренить злостных вредителей.
Полезные сорняки на участке
Не все сорняки – вредители, в саду и на огороде можно встретить и полезные дикорастущие растения.
Василек синий – целебная приправа в кулинарии. Считается, что растение обладает мочегонным, болеутоляющим, желчегонным и ранозаживляющим действием. Настой из цветков помогает при фурункулах, экземе и коньюктивите. Высушенные соцветия василька используются как натуральный краситель при приготовлении блюд.
Луговому клеверу приписывают антисептические и противовоспалительные качества. Растение используют при лечении атеросклероза и сухого кашля. Из цветков клевера и молодых листьев готовят весенние салаты, а засушенные побеги добавляют во время варки вторых блюд.
Молодая крапива – ценный подарок весны. Она содержит большое количество витамина С и каротина. Употребление отвара из крапивы стимулирует обмен веществ в организме и способствует снижению веса. Крапиву используют при варке супов, борщей, приготовлении мяса, омлетов и других блюд.
Хвощ полевой – многолетнее растение в высоту до 60 см. Применяется при лечении мочевого пузыря и как противомикробное средство. Сухую траву заваривают, настаивают и пьют перед едой. Для улучшения кровообращения и облегчения состояний при ревматических заболеваниях рекомендуют принимать «хвощевые» ванны.
Горец птичий (спорыш) – однолетний сорняк, растущий в садах, парках, на огородах и газонах. Спорыш содержит много биологически активных веществ: кумарины, флавоноиды, эфирные масла, витамины и фенолкарбоновые кислоты. Растение может использоваться как жаропонижающее средство при простудных заболеваниях или наружно – для ускорения заживления ран и язв.
Профилактика появления сорняков
Бороться с сорняками на огороде и газоне будет гораздо легче, если их будет не слишком много. Для этого надо проводить профилактические мероприятия.
Рекомендации по уходу за газоном, снижающие вероятность появления сорняков:
- Газон не следует подстригать слишком коротко. Молодую лужайку надо стричь понемногу, но часто.
- Регулярный полив в засушливую погоду. Газон не должен поредеть после лета, иначе пустующие участки быстро займут сорняки.
- Чистка газона граблями препятствует росту ползучих сорняков.
- Подкормка поможет окрепнуть газонной траве и не даст возможность сорнякам вытеснить ее.
- Выявление болезней и вредителей. Если не удалось защитить газон, то поредевший участок надо засеять свежими семенами трав до появления на нем сорняков.
- Любое сорное растение надо сразу же удалить, пока оно не отрастило объемные корни и не созрело до семян.
Профилактика появления огородных сорняков:
- На участке надо выделить огражденное место под компостную яму, куда в течение сезона будут складываться все остатки сорняков с огорода.
- Осенью, после сбора урожая, надо перекопать огород, не разбивая комки. На поверхности окажутся многие корневища сорняков и за зиму они вымерзнут.
- Ранней весной, перед посадкой овощных культур, проводят повторную перекопку и стараются максимально удалить оставшиеся корни растений.

Виды сорняков: название, описание, фото
Один из наиболее важных пунктов по уходу за огородными грядками, садом и газоном – профилактика и удаление сорняков. Способы борьбы зависят от вида сорного растения, его биологических особенностей и места произрастания. Рассмотрим основные типы сорняков и опишем методы контроля над их распространением.
Содержание
- Вред и польза от сорных растений
- Классификация сорняков
- Cорные растения на огороде: названия, описания, фото
- Газонные сорняки: названия, описания, фото
- Полезные сорняки на участке
- Профилактика появления сорняков
Вред и польза от сорных растений
Сорняками принято называть растения, которые «поселились» на участке помимо возделываемых культур. Борьба с ними отнимает у дачников очень много времени и сил. На сегодняшний день известно более 2000 сорных растений, в том числе ядовитых и вредных для животных – около 100 наименований.
Борьба с ними отнимает у дачников очень много времени и сил. На сегодняшний день известно более 2000 сорных растений, в том числе ядовитых и вредных для животных – около 100 наименований.
Как бы мы ни старались, но избавиться один раз и навсегда от сорных растений не получиться – они попадают на участок через самые разные источники:
- некоторые семена сорняков «сидят» в почве и ожидают благоприятных условий для прорастания;
- органические удобрения – если компост не прошел должную обработку;
- некачественный посевной материал;
- семена заносятся ветром, животными, людьми (на подошве обуви).
Сорные растения причиняют ощутимый вред земледелию и ландшафтному дизайну:
- заглушают культурные растения и снижают их урожайность;
- выделяют в почву вредные вещества;
- поглощают большое количество питательных веществ и воды;
- создают тень;
- могут стать причиной отравления домашних животных;
- являются очагом для развития болезней и вредителей с/х культур.

Но, не все так однозначно. Агротехники отмечают, что некоторые виды сорных растений приносят и пользу. Сорняки с мощными корнями разбивают уплотнения грунта и разрыхляют почву, извлекают полезные вещества из большой глубины, которая не доступна газонной траве и некоторым огородным культурам. Из таких растений получается хорошее удобрение.
Классификация сорняков
Все сорные растения классифицируют по трем основным биологическим признакам:
- продолжительность жизни;
- способ размножения;
- способ питания.
Зависимо от продолжительности жизни сорняки подразделяют на малолетние и многолетние виды.
Малолетние «бурьяны» размножаются семенами. Эта группа включает в себя:
- эфемеры – вегетационный период меньше одного сезона;
- яровые – вегетационный период такой же, как и у однолетних огородных растений; чаще остальных сорняков засоряют культурные посевы;
- озимые однолетние – всходят в начале осени; засоряют посадки многолетних трав и посевы пшеницы;
- двулетние – полный цикл развития включает два вегетационных периода.

Многолетние сорняки могут расти на одном месте до 4-х лет. После того, как семена созревают, наземные органы растения отмирают, а корневая система продолжает развиваться. Ежегодно от подземной части отрастают новые стебли. Многолетники размножаются вегетативно или семенами.
По способу питания различают такие виды сорняков:
- Непаразитные сорняки имеют самостоятельный тип питания и их развитие не зависит от других растений. Наиболее многочисленная группа.
- Полупаразитные сорняки имеют способность к фотосинтезу, но в тоже время, частично питаются за счет других культур – присасываются к наземным органам или корням растений. Типичные представители: ремнецветник европейский, омела белая, марьянник полевой, мытник болотный, зубчатка поздняя.
- Паразитные сорняки – растение не имеет зеленых листьев, корней, не способно к фотосинтезу и питается исключительно за счет другого растения. Сорняки-паразиты крепятся к корням или стеблям культурных растений. К этой группе относятся: повилика клеверная, повилика льняная, заразиха подсолнечниковая. Способ размножения – семенами.
 К этой группе относятся: повилика клеверная, повилика льняная, заразиха подсолнечниковая. Способ размножения – семенами.
К этой группе относятся: повилика клеверная, повилика льняная, заразиха подсолнечниковая. Способ размножения – семенами. Сорные растения на огороде: названия, описания, фото
Пырей ползучий обитает на полях, огородах, в садах, в поймах рек, вдоль дорог. Имеет глубокую корневую систему и быстро распространяется по участку. Стебель сорняка – прямостоячий, листья – плоские, длинные с шероховатой поверхностью. Пырей очень устойчив к неблагоприятным условиям, размножается корневищами практически на любом типе почве, многолетнее растение семейства злаковых. От пырея надо избавляться, особенно если планируется посадка картофеля.
Если не бороться с пыреем, газон может полностью зарости сорняками: фото
Вьюнок полевой (березка) – многолетний сорняк, обволакивающий стебли растений. Одно растение способно запутать до 2-х квадратных метров площади посевов. Больше всего от березки страдают ягодные кустарники. Длина стебля – до 180 см, листья – стреловидные, плод – двухгнездная коробочка. Разветвленные корни полевого вьюнка уходят в землю на глубину до пяти метров, поэтому полностью избавиться от растения очень сложно – корни придется выкапывать.
Больше всего от березки страдают ягодные кустарники. Длина стебля – до 180 см, листья – стреловидные, плод – двухгнездная коробочка. Разветвленные корни полевого вьюнка уходят в землю на глубину до пяти метров, поэтому полностью избавиться от растения очень сложно – корни придется выкапывать.
Портулак огородный – однолетнее растение с красноватым толстым стеблем и мясистыми листьями. Длина стебля – около 60 см, побеги одного растения могут покрыть значительную площадь огорода. Портулак может использоваться в лекарственных целях и в кулинарии.
Мокрица (звездчатка) – зимующий однолетник, эфемер. Всходы сорняка появляются ранней весной, а когда наступает время для прорастания культурных растений – мокрица разрастается в сплошной зеленый ковер. От этого сорняка сильно страдают посевы моркови.
В старину по мокрице предсказывали погоду на ближайшее время. Считалось, что если после восхода солнца цветки звездчатки не поднимались и не раскрывались, то днем следует ждать осадков
Щирица запрокинутая – яровый ранний однолетник, отличающийся очень высокой плодовитостью. Семеня сорняка не боятся механических воздействий и сохраняют способность к прорастанию на протяжении 5-40 лет. Растение имеет высокий опушенный стебель (до 150 см), яйцевидно-ромбические листья, цветки собраны в густое метельчатое соцветие. Семена могут прорасти с глубины не больше 3 см.
Семеня сорняка не боятся механических воздействий и сохраняют способность к прорастанию на протяжении 5-40 лет. Растение имеет высокий опушенный стебель (до 150 см), яйцевидно-ромбические листья, цветки собраны в густое метельчатое соцветие. Семена могут прорасти с глубины не больше 3 см.
Ежовник (куриное просо) – засоряет овощные культуры (подсолнечник, морковь, свеклу) в начале их роста. В дождливые года способен полностью заглушить изреженные молодые посевы. Стебель достигает 120 см в высоту, листья – широколинейные, заостренные по краям, соцветие – метелка с колючими одноцветковыми колосками, наполненная семенами, которые похожи на просо.
Подмаренник цепкий – ранний яровой однолетник. Отличительная особенность – ствол и листья сорняка буквально цепляются за одежду. Подмаренник цепкий появляется на плодородных, богатых известняком почвах. Коневая система сорняка – стержневая, стебель – четырехгранный высотою до 1 м, на ребрах стебля есть маленькие, загнутые вниз шипы. Цветки собраны в густые метелки, период цветения – лето.
Цветки собраны в густые метелки, период цветения – лето.
Пастушья сумка – зимующий однолетник, цветущий с весны до поздней осени. Жизнеспособность семян сохраняется почти 35 лет. Высота стебля около 20-40 см, листья перисто-рассеченной формы, корень – стержневой. Белые мелкие цветки собраны на самой верхушке стебля, цветение сорняка продолжается все лето. За это время растение дает 2-4 поколения (семена опадают и сразу же прорастают).
Ярутка полевая – однолетнее растение с характерными округлыми плодами с вырезом на верхушке. Высота стебля – не более 40 см, листья собраны в розетку и находятся у земли. За период цветения ярутка дает до 50 тысяч семян. Максимальная глубина, с которой семена могут прорости – 5 см.
Осот розовый, более известный, как бодяк полевой – распространенное многолетнее растение сорняк, засоряющее любые посевы. Высота осота может достигать 1,5 м, поверхность стебля колючая. Форма листа ланцетная, по краям имеются колючки.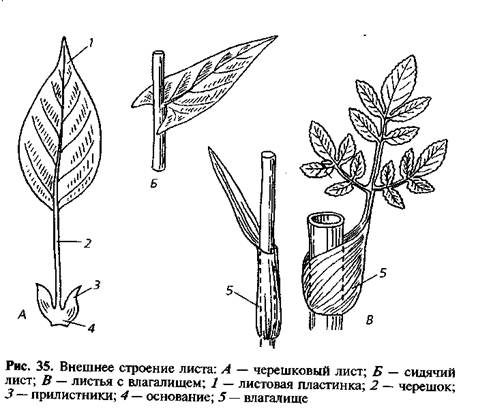 Взрослое растение имеет мощную корневую систему, прорастающую вглубь до 6 метров. На изгибах корня закладываются почки, дающие развитие надземным побегам.
Взрослое растение имеет мощную корневую систему, прорастающую вглубь до 6 метров. На изгибах корня закладываются почки, дающие развитие надземным побегам.
Основной источник засорения огорода бодяком – вертикальный корень. При борьбе с осотом необходимо уничтожать корневую систему, расположенную на глубине до 60-70 см
Газонные сорняки: названия, описания, фото
Мятлик – однолетний сорняк. В начале роста растение незаметно, но со временем сорняк зацветает и выделяется некрасивыми пятнами на газонном ковре. Мятлик хорошо себя чувствует на уплотненных почвах в низинах. Если своевременно вывести сорняк, то он долго не будет появляться на газоне.
Не избежать появления на газоне одуванчиков, семена которых разносятся ветром. Лучше всего одуванчики «приживаются» на молодых, редко засаженных газонах. Сорное растение имеет мясистые корни, в которых содержится большой запас питательных веществ. Это надо учитывать при борьбе с сорняком – для полного уничтожения понадобится не одна обработка гербицидами избирательного действия.
Лютик ползучий – многолетний сорняк со стелющимся по земле стеблем. Длина растения – около 1 м, листья имеют тройчатую форму. Лютик размножается семенами и вегетативно. Благоприятно развивается во влажных и недостаточно освещенных местах.
Мох, как и лютик, появляется на сырых почвах в низинах. Для борьбы с ним газон надо регулярно аэрировать и делать дренажные канавы. Высота мха – не более 50 см. Если не бороться с этим сорняком, то он способен полностью вытеснить газонные злаки и привести к заболачиванию почвы.
Появление мха может свидетельствовать о «бедном» составе почвы, нехватке питательных элементов и чрезмерном уровне кислотности грунта
Подорожник часто растет на переуплотненной, вытоптанной почве или в местах, где наблюдается застой влаги. Для удаления взрослых растений подойдет специальная садовая вилка. Если подорожники значительно разрослись по газону, то надо использовать гербициды избирательного действия.
Вероника нитчатая селится на увлажненной почве, богатой на полезные элементы. Высота растения – не более 12 см, стебель тонкий, цветы нежно лилового цвета. Сорняк размножается побегами.
Клевер – наиболее проблемный сорняк, доставляющий массу хлопот владельцам газонов. Многолетнее растение высотою 15-50 см, корневая система – стержневая, листья яйцевидной формы. Появление клевера может сигнализировать о нехватке в почве азота.
Активный рост клевера могут спровоцировать калийные и фосфорные удобрения, внесенные весной
Марь белая — морозоустойчивый сорняк на участке, растущий с ранней весны до глубокой осени. Высота растения может достигать 1,5 м. Максимальная глубина, с которой семена могут прорости – 10 см. Растение надо удалять до цветения, так как один сорняк может дать около 500 тысяч семян с различными сроками всходов.
Кислица обыкновенная (заячья капуста)– многолетнее растение с мощной корневой системой. Сорняк растет группами и с легкостью «забивает» газонную траву. Кислица очень устойчива к многим химическим препаратам, поэтому наиболее действенный способ – вырывание сорняка с корнем.
Представленные фото и названия сорняков помогут вовремя распознать и искоренить злостных вредителей.
Полезные сорняки на участке
Не все сорняки – вредители, в саду и на огороде можно встретить и полезные дикорастущие растения.
Василек синий – целебная приправа в кулинарии. Считается, что растение обладает мочегонным, болеутоляющим, желчегонным и ранозаживляющим действием. Настой из цветков помогает при фурункулах, экземе и коньюктивите. Высушенные соцветия василька используются как натуральный краситель при приготовлении блюд.
Луговому клеверу приписывают антисептические и противовоспалительные качества. Растение используют при лечении атеросклероза и сухого кашля. Из цветков клевера и молодых листьев готовят весенние салаты, а засушенные побеги добавляют во время варки вторых блюд.
Молодая крапива – ценный подарок весны. Она содержит большое количество витамина С и каротина. Употребление отвара из крапивы стимулирует обмен веществ в организме и способствует снижению веса. Крапиву используют при варке супов, борщей, приготовлении мяса, омлетов и других блюд.
Хвощ полевой – многолетнее растение в высоту до 60 см. Применяется при лечении мочевого пузыря и как противомикробное средство. Сухую траву заваривают, настаивают и пьют перед едой. Для улучшения кровообращения и облегчения состояний при ревматических заболеваниях рекомендуют принимать «хвощевые» ванны.
Горец птичий (спорыш) – однолетний сорняк, растущий в садах, парках, на огородах и газонах. Спорыш содержит много биологически активных веществ: кумарины, флавоноиды, эфирные масла, витамины и фенолкарбоновые кислоты. Растение может использоваться как жаропонижающее средство при простудных заболеваниях или наружно – для ускорения заживления ран и язв.
Профилактика появления сорняков
Бороться с сорняками на огороде и газоне будет гораздо легче, если их будет не слишком много. Для этого надо проводить профилактические мероприятия.
Рекомендации по уходу за газоном, снижающие вероятность появления сорняков:
- Газон не следует подстригать слишком коротко. Молодую лужайку надо стричь понемногу, но часто.
- Регулярный полив в засушливую погоду. Газон не должен поредеть после лета, иначе пустующие участки быстро займут сорняки.
- Чистка газона граблями препятствует росту ползучих сорняков.
- Подкормка поможет окрепнуть газонной траве и не даст возможность сорнякам вытеснить ее.
- Выявление болезней и вредителей. Если не удалось защитить газон, то поредевший участок надо засеять свежими семенами трав до появления на нем сорняков.
- Любое сорное растение надо сразу же удалить, пока оно не отрастило объемные корни и не созрело до семян.
Профилактика появления огородных сорняков:
- На участке надо выделить огражденное место под компостную яму, куда в течение сезона будут складываться все остатки сорняков с огорода.
- Осенью, после сбора урожая, надо перекопать огород, не разбивая комки. На поверхности окажутся многие корневища сорняков и за зиму они вымерзнут.
- Ранней весной, перед посадкой овощных культур, проводят повторную перекопку и стараются максимально удалить оставшиеся корни растений.
Карантинные сорные растения / Россельхознадзор
Карантинные сорные растения — это особо вредные виды сорняков, не произрастающие или ограниченно распространённые на территории РФ или отдельного региона. Степень агрессивности этих сорняков представляет значимую угрозу огородным и полевым культурам, а также человеку и животным.
Они забирают значительную часть питательных веществ и влаги, из-за них снижается урожайность с/х культур, растения вытесняются со своего места произрастания. Зерновые, собранные с зараженных полей, отличаются плохим качеством. Появление агрессивных растений приводит к изменению структуры видового сообщества. Еще один фактор, влияющий на развитие с/х и огородных культур, – распространение патогенной микрофлоры, вредителей и болезней. При низкой агротехнике сорняки губят почти весь урожай. Некоторые сорта карантинных растений-сорняков очень токсичны для животных. При выпасе на полях, заселенных подобными растениями, происходит их отравление. Основным вредом для людей считаются аллергические проявления, которые могут быть разной степени тяжести.
Кроме перечисленных проблем, карантинные организмы приводят к снижению качества убранного урожая, способны вызвать поломку уборочной техники и дополнительные расходы. Дополнительно приходится очищать урожай от семян сорных трав.
Не все карантинные виды сорняков произрастают в РФ. Некоторые отсутствуют полностью, другие распространены ограниченно. Но есть такие, которые растут почти на всей территории России и представляют значительную опасность. Это повилики; горчак ползучий; амброзии: полыннолистная, трехраздельная, многолетняя; паслены: колючий, трехцветковый.
Повилика.
На территории России растут более 30 видов повилики. Самыми опасными и распространенными являются повилики: полевая, клеверная, хмелевидная, льняная, короткоцветковая перечная и повилика лемана. Они крайне плодовиты и жизнеспособны.
У повилики отсутствует корневая система. Питаются повилики, прикрепляясь присосками к растению-хозяину. Присоски (гаустории) глубоко проникают в ткань культурного растения. Стебли сорняка почти лишены хлорофилла, нитевидные, вьющиеся, покрыты листочками-чешуйками и мелкими многочисленными цветочками. В качестве растения — хозяина повилики используют многолетние и однолетние травы, деревья, кустарники, виноградники. Высасывая все соки, повилики нарушают обмен веществ «хозяина», растение ослаблено, отстает в росте и развитии. Нередко целые массивы культурных растений, зараженных паразитом, погибают. Сено, полученное из зараженных сорняком трав, утрачивает питательные свойства, быстро плесневеет, способствует болезням и гибели животных. Повилики, внедряясь внутрь растений, переносят вирусные болезни.
Распространение повилики происходит в основном с семенами культурных видов при их недостаточной очистке. Если в корм скоту попало сено, зараженное повиликой, то семена сорняка попадут с навозом на удобряемые им площади. Семена повилики сходны с семенами растений-хозяев, это результат паразитической адаптации, и он усложняет очистку семян культурных трав.
Горчак ползучий (розовый).
Этот многолетний корнеотпрысковый сорняк растет в виноградниках и садах, на пастбищах и лугах, вдоль шоссе, на железнодорожных откосах и пустующих землях. Растение обладает мощной корневой системой, его главный вертикальный корень проникает в почву более, чем на десятиметровую глубину и имеет разветвленную систему горизонтальных корней. Размножается как семенами (их всхожесть в почве составляет 3-5 лет), так и корневищами. Распространяется горчак с плохо очищенным посевным материалом, с соломой и сеном. Горчак чрезвычайно ядовит для лошадей, у коров вызывает горечь молока. Корневая система горчака очень быстро разрастается, лишая культурные растения влаги и минералов и снижая их урожайность вдвое. Одно растение сорняка в течение года, разрастается в куртину 5-6 м диаметром, а его переплетенные корни не дают никаких шансов культурным растениям. Светолюбивый горчак не образует семян в затененной местности, рост его корневой системы замедляется, но сохраняет способность к взрывному росту при наступлении благоприятных условий.
Амброзия полыннолистная.
Мощные корни и массивная надземная часть сорняка угнетают культурные растения. В ходе вегетационного периода, амброзия буквально высасывает из почвы воду и минеральные вещества, иссушает и истощает почву, затеняет культурные растения, вытесняя их с полей и пастбищ. В период своего цветения, сорняк выделяет огромную массу пыльцы, вызывающую тяжелые аллергические реакции. Цветение амброзии продолжается с июля до октября. Однолетнее растение достигает высоты 1,8 м, стержневой корень уходит вглубь почвы на 4 метра. Размножается амброзия семенами, количество которых с одного куста может достигать 40 тысяч. Высокая всхожесть наблюдается и у невызревших семян сорняка. Растение хорошо адаптировано к затоплению и частому скашиванию.
Амброзия трехраздельная.
Однолетнее растение, яровой сорняк крупного размера, его стебель деревенеет к осени. Имеет много сходных свойств с амброзией полыннолистной, отличаясь ранним вызреванием, более крупным размерам семян и их плавучестью, в силу чего начало заражения происходит в низинных, подтопляемых местностях. Распространение, вред и методы борьбы с этим сорняком такие же, но этот вид сорняка лучше уничтожается химическими средствами, т.к. поверхность листа у него больше.
Амброзия многолетняя.
Многолетний, корнеотпрысковый сорняк. Главный корень растения стержневой, имеет многочисленные отростки, из которых вырастают новые надземные побеги. Часто встречается на пастбищах, лугах. Не вытесняется многолетними травами, корневища растений устойчивы к морозам. Этот сорняк трудно искоренить, вызывает снижение урожайности и качества культурных растений, ослабляет продуктивность пастбищ, т.к. этот сорняк не употребляется в пищу скотом. Пыльца многолетней амброзии сильнейший аллерген.
Паслен колючий.
Все части растения густо усажены шипами. Диаметр одного растения порядка 70 см, плодоносит с августа по октябрь. На каждом растении вызревает около 180 ягод, в каждой из которой, находится от 50 до 120 семян, которые приобретают всхожесть после зимовки и сохраняют ее 7-10 лет. После вызревания семян, растение обламывается и перекатывается на большие расстояния. Семена сорняка разносятся ветром, перемещаются на колесах автотранспорта. Главный корень растения прорастает в почву на глубину 3 метра. При благоприятных условиях, наземная часть сорняка наращивает огромную зеленую массу высотой около метра. Произрастает на обочинах, пустырях, вытесняя оттуда все прочие травы. Недостаток света в начале вегетации растения действует на него угнетающе. Засоряет посевы, пастбища, огороды и сады. Разветвленная корневая система сорняка лишает культурные растения питания и воды. Потери урожая на зараженных участках составляет 40-50%. Листья паслена ядовиты, а колючки повреждают ЖКТ животных и ранят ротовую полость. Паслен служит растением — хозяином для картофельной моли, колорадского жука и некоторых вирусов.
Паслен трехцветковый.
Однолетник с высокой семенной продуктивностью. На одном растении вызревает 10-14 тысяч семян, а их всхожесть сохраняется в течение 9 лет. Образует крупные кусты с жесткими ветвями. Этот сорняк способен давать придаточные корни от стеблей и легко укореняться. Удаленное прополкой растение, оставленное на земле, с легкостью укореняется заново. Семена паслена клейкие, поэтому прилипают к различным предметам и разносятся на большие расстояния. Сорняк ядовит, имеет неприятный трупный запах.
Основные способы противодействия распространению карантинных сорных растений это профилактические меры: очистка посевного материала и фуражного зерна, переработка которого должна осуществляться по технологии, лишающей семена всхожести. Внесение в почву перепревшего навоза, в котором все семена сорняков утратили жизнестойкость.
Соблюдение севооборота, зяблевая вспашка, плоскорезное возделывание почвы, уход за посевами, боронование, междурядные обработки пропашных культур, предотвращение повторного обсеменения почвы сорняком. Сочетая агротехнические способы с применением разрешенных на территории РФ гербицидов, можно сдержать распространение карантинных сорняков.
Источник: Управление Федеральной службы по ветеринарному и фитосанитарному надзору по Томской области
Двудольные сорняки: описание, меры борьбы
Растения, не культивируемые человеком, но сопровождающие посадки сельскохозяйственных культур, относятся к сорным растениям или сорнякам. Некоторые из них попадают на сельскохозяйственные земли из окружающей среды, с посевным материалом, разносятся животными и птицами.
Все сорные растения негативно влияют на количество и качество урожая, снижая его на 10 — 50 %. Сорняки бывают однолетними, двулетними, многолетними. По строению семени покрытосеменных растений различают однодольные и двудольные сорняки. Наличие двух семенных долей у представителей класса двудольных, обеспечивает их семя большим запасом питательных веществ, что в свою очередь дает более мощный рост как корню, так и наземной части стебля.
При прорастании семян, двудольные дают стебель с двумя зародышевыми листочками (по количеству долей). Настоящий лист начинает формироваться из почки, расположенной между ними. У однодольных, как правило, сразу выходят настоящие листья. Так же большинство двудольных сорняков имеют мощный стержневой корень, уходящий на достаточную глубину. Что бы своевременно провести агротехнические мероприятия по борьбе с сорными растениями класса двудольных, рассмотрим типичных представителей, характерных для большинства районов.
Содержание:
Однолетние и двулетние двудольные сорняки
- Василек синий — сорняк со стеблем высотой до 90 см, прорастает в мае, зацветает в июне — сентябре. Цветки у василька — голубых, синих, фиолетовых оттенков. Семена очень мелкие, вызревают в большом, до 1500 с одного цветка, количестве. Всхожесть в сухом состоянии семена сохраняют от 3 до 10 лет. Засоряет василек синий, как правило посевы злаковых культур.
- Горец почечуйный — мощное растение более метра в высоту. Цветки розовые, зеленовато — розовые, собранные в длинные кисти. Горец отличается еще большей плодовитостью, дает до 3000 семян с максимальной жизнеспособностью до 10 лет. Сопровождает посадки овощных, масленичных, бобовых растений.
- Молочай — солнцегляд — крепкое растение высотой до полуметра, с удлиненными листьями и мощным хорошо развитым корнем. Прорастает в мае, цветет все лето. Низкая всхожесть семян компенсируется их огромным количеством. Предпочитает расти среди зерновых, бобовых, кормовых культур.
Также к повсеместно распространенным однолетним двудольным сорнякам относятся:
- марь
- незабудка
- осот огородный
- паслен
- белена
- пастушья сумка
Многолетние двудольные сорняки
Одуванчик лекарственный — повсеместно распространенное растение, удлиненные листья собраны в прикорневую розетку, цветы имеет ярко — желтые, семена легко разносятся ветром. Встречается в огородных и кормовых культурах.
Полынь — мощное высокорослое, напоминающее кустарник растение. Прорастает в мае, цветет в июле — августе. Способна прорасти с глубины до 10 см. Дает несколько десятков тысяч семян. Засоряет зерновые, бобовые, кормовые, огородные культуры.
Молочай лозный — сорняк, размножающейся не только семенами, но и корневыми отпрысками. Даже небольшой остаток корня дает возможность разрастания молочая на всей территории участка. Сопровождает практически все огородные культуры.
Так же к этой группе сорняков относятся:
- подорожник большой
- осот полевой
- горошек мышиный
- клевер ползучий
- лютик полевой
- лютик ползучий
Меры борьбы с двудольными сорняками
Механические меры — заключаются в удалении сорняков, их подземных и наземных частей. Сделать это можно как в ручную, на небольших участках, так и при помощи ручных орудий труда:
- тяпок
- корнеудалителей
- плоскорезов
На площадях до одного гектара целесообразно использовать для прополки мотоблоки со
специальным навесным оборудованием. Укрытие междурядий светонепроницаемым материалом значительно снизит количество сорняков на участке.
Химические методы — обработка сельскохозяйственных земель гербицидами, губительными для двудольных сорняков. Почвенными гербицидами земля обрабатывается до посева, либо до всходов семян культурных растений или осенью, после уборки урожая. Проникая на глубину в несколько сантиметров, они уничтожают семена сорных растений до их прорастания. Против двудольных сорняков используют следующие препараты:
- Пионер
- Комманд
Послевсходовые гербициды можно разделить на гербициды сплошного действия и избирательного действия. Химикаты сплошного действия уничтожают, как правило, и сорняки и культурные растения. К ним относятся:
- Раундап
- Ураган
- Снайпер
- Лонтрел 300
- Лазурит СП
Стоит сказать, что при соблюдении рекомендованных концентраций и норм внесения, современные гербициды не наносят вреда здоровью человека. Иногда бороться с двудольными сорняками только механическими методами бывает очень сложно, ведь их семена не теряют всхожести десяток лет, но выбор методов борьбы с ними остается за огородником.
Видео о борьбе с сорняками на участке:
Карантинные сорные растения | Министерство сельского хозяйства и рыбной промышленности Астраханской области
Карантинные сорные растения занимают особое положение среди большого количества сорных трав из-за высокой вредоносности. Будучи занесёнными из других ботанико-географических областей, они акклиматизируются на новой территории и начинают активно размножаться, т.к. на новом месте они оказываются вне досягаемости для вредителей и болезней, которые повреждали их на родине и сдерживали их развитие. В отсутствие сдерживающих факторов такие растения становятся особенно злостными сорняками.
При засорении полей карантинными сорняками наблюдается значительное снижение урожайности культурных растений — а при низкой агротехнике и полное уничтожение сельхозкультур. Многие карантинные сорные растения являются переносчиками возбудителей вирусных и бактериальных заболеваний, опасных для культурных растений. Большинство карантинных сорняков не поедаются животными, поэтому засорённые ими луга и пастбища становятся малопродуктивными. Карантинные сорняки, имеющие грубые скелетные ветви (амброзия полыннолистная, амброзия трёхраздельная) забивают режущие органы с/х техники, в результате чего снижается качество и производительность уборки, увеличиваются расходы на дополнительную очистку урожая с/х культур. Солома, убранная с полей, засорённых паслёном колючим и ценхрусом малоцветковым, становится непригодной ни для скармливания животным, ни для подстилки из-за наличия колючек. Попадая в полость рта животных, колючки и колючие колоски вызывают язвы и опухоли. Корма, содержащие растения повилики, могут стать причиной отравления животных, а зелёные корма, содержащие амброзию полыннолистную, делают вкус молока горьким. Также хорошо известно, что кроме вреда животным и ущерба животноводству, пыльца растений из рода амброзия является сильным аллергеном и вызывает у людей заболевание поллинозом. Лечение аллергии, вызванной пыльцой амброзии, продолжительное и не всегда успешное. Интенсивное расселение карантинных сорных растений может привести к нарушению стабильности флоры региона, что, в свою очередь, может быть вредно для животного мира и может привести к дестабилизации и разбалансировке всей экосистемы.
Помните, что если карантинные виды сорняков «поселяться» в вашем огороде, справиться с ними самостоятельно вы не сможете, так как большинство из них являются трудноискоренимыми видами. Эти пришлые растения не только заберут всю воду и питательные элементы из почвы, но могут полностью лишить вас урожая, а некоторые из карантинныхсорняков ядовиты для человека и животных (паслен трехцветковый, паслен колючий).
Обратите внимание! Многие карантинные сорные растения по внешнему виду схожи с обычными сорняками. Так, амброзию можно принять за обычную полынь, горчак спутать с часто встречающимся у нас осотом розовым, а паслен трехцветковый и вовсе не отличить от обычного паслена черного.
Основными элементами системы мероприятий, направленных на борьбу с карантинными сорными растениями являются:
— карантинный фитосанитарный контроль растительной продукции, транспортных средств (как при ввозе на территорию РФ, так и при внутрироссийских перевозках)
— введение карантинных ограничений по использованию засорённой продукции;
— обследование земельных угодий на выявление первичных очагов карантинных сорных растений, мониторинг территории;
— организация работы по борьбе с карантинными сорняками, принятие мер по ликвидации первичных очагов.
Владельцам и пользователям подкарантинных объектов различных форм собственности хотелось бы напомнить: действующее законодательство предусматривает проведение систематических карантинных фитосанитарных обследований, направленных на своевременное выявление карантинных объектов и предупреждение их распространения.
Тема занятия «Сорные растения и их внешний вид».
Тема занятия: «Сорные растения и их внешний вид».
Цель занятия: сформировать понятие о сорных растениях как обязательной части растительности и роли сорняков в сельском хозяйстве.
Задачи:
Образовательные:
— познакомить учащихся с многообразием сорных растений.
— рассказать о причинах снижения урожайности сельскохозяйственных культур.
Развивающие:
— создать условия для развития умственных, коммуникативных способностей учащихся.
— способствовать развитию памяти, речи и технологического мышления в ходе определения способов борьбы с сорняками в посевах сельскохозяйственных культур.
Воспитательные:
— способствовать воспитанию таких качеств личности как трудолюбие, дисциплинированность, аккуратность, точность.
— подвести учащихся к пониманию того, что от уровня знаний зависит будущий урожай.
Методы: объяснительно — иллюстративный, частично- поисковый, наглядные.
Оборудование: гербарные образцы, фотографии.
Ход занятия.
I. Изучение нового материала.
1. Понятие о сорных растениях (рассказ).
Сорняки растут не везде, а только там, где они не нужны. (М. Генин)
Сорными называют такие растения, которые не возделываются человеком, но засоряют посевы сельскохозяйственных культур и другие угодья. Засорители – растения, относящиеся к культурным видам, но не возделываемые на данном поле и засоряющие посевы основной культуры.
На территории России произрастает около 1500 видов сорных растений из них 400видов ядовитые. Праздник сорняков 19 июня- Пропольник. В этот день сорняки не проклинают, а почитают как магические атрибуты Матушки-Земли. Ведь недаром сорняки красуются на картинах известных художников. В Русском музее и Третьяковской галерее хранятся два этюда художника И.И. Шишкина, посвященные растению-сорняку. Один из них называется «Сныть-трава».
Понятие «сорные растения» относительно, зависит от места произрастания растения. Примеры относительности: взять, к примеру, льнянку обыкновенную. В народе растение это зовут львиным зевом. Бороться с ней трудно, она очень быстро размножается корневыми отпрысками. Но в то же время это — лекарственное растение, нужное людям. И пчелы любят посещать ее (она одно из медоносных растений).
2. Вред сорных растений.
Работая в парах, дайте ответ на вопрос: «В чем заключается «вред» сорных растений?
Снижается урожайность и ухудшается качество продукции. Конкуренция с культурными растениями за свет, воду, питательные вещества.
На сильно засорённых участках может быть снижение урожая в 1,5-2 раза
Корни сорняков более развиты, чем у культурных растений (у осота, хвоща корни проникают до глубины 5,5-7,5 м). Полынь поглощает воды в 2 раза больше, чем пшеница.
Потребление элементов питания сорняками значительно более интенсивное. Хвощ полевой – потребляет в 5-7 раз больше, чем озимая пшеница, следовательно, при высокой засорённости большая часть удобрений может уходить на “прокорм” сорняков.
На сорняках размножаются вредители и сохраняются возбудители болезней.
Затенение от сорняков снижает эффективность фотосинтеза культурных растений, уменьшает содержание белка в зерне. Ухудшается качество продукции. Многие сорняки и их семена ядовиты. Хвощ, лютик в сене вызывают отравление животных. Некоторые сорняки вызывает аллергию (амброзия).
Совместно формируем вывод об отрицательной роли сорных растений:
1. Угнетение жизнедеятельности выращиваемых растений.
2. Снижение качества урожая.
3. Истощение почв.
4. Заросли сорняков – первичные резерваты вредителей и болезней с/х растений.
Ликвидация сорных растений – первейшая задача в обеспечении высокой культуры земледелия.
3. Биологические особенности сорных растений. (рассказ, беседа)
1) Плодовитость. С 1 м2 осота жёлтого семян хватит для того, чтобы засеять 10 га
2) Разнообразные способы распространения. Семена сорняков похожи по форме и размеру с семенами культурных растений, поэтому тяжело отделить при сортировке.
3) Высокая жизнеспособность семян. Семена сорняков, оставаясь в почве, много лет способны сохранять всхожесть.
4) Растянутое прорастание.
5) Способность размножаться вегетативно. У пырея на 1 м2 длина корневищ до 500 м, их масса -2,9 кг/м2; обломки корней дают новые растения.
6) Раннее созревание. Семена осыпаются до начала уборки; недозрелые семена сохраняют всхожесть
4. Типы сорных растений.
Демонстрация гербария. Практическая работа в группах по теме. Цель работы: определить и описать наиболее распространенные сорные растения. Данные зафиксировать в рабочей тетради.
Физминутка.
5. Классификация сорняков.
а) по способу питания б) по продолжительности жизни в) по способу размножения.
По способу питания: паразиты, непаразиты (фотосинтезирующие) – наиболее распространены, полупаразиты.
По продолжительности жизни: малолетние, двулетники – цикл развития – 2 года (донник жёлтый, чертополох), многолетние – живущие более 2 лет, размножаются семенами и вегетативно.
6. Пословицы и поговорки про сорняки.
Дай волю осоту, и огурцов на белом свете не станет.
Где сорняк цветёт, там хлеб вянет.
Сорняки в поле оставили, урожай убавили.
Афоризмы и цитаты про сорняки.
Что такое сорняк? Растение, польза которого еще не открыта. (Ральф Эмерсон)
Они знают, это факт, где расти, как нас одурачить, как замаскироваться под самое полезное растение, и поэтому я заключаю, что у сорняков есть мозги. (Дайанн Бенсон)
Сорняк (басня в стихах).
Среди собратьев на песочной куче
Сорняк ютился тощий и колючий.
Томимый жаждой, в вечной тесноте
Он ненавидел все растенья те,
Что наливались соком на свободе
На клумбе и на грядках в огороде:
«Меня тут тля грызет, и солнце жжет,
А их от бед хозяйка бережет
От тли опрыснет, грунт взрыхлит, прополет,
Польет из лейки теплою водою…
А я весь высох, из песка торчу!
Я тоже в огороде жить хочу!»
Но жук ему сказал: «Не думай даже!
Ведь каждый там не просто так посажен.
Взгляни-ка, друг, еще раз за забор:
Вот роза, чтоб пленять хозяйский взор,
А вот клубника, чтоб зимой в варенье
Дарить хозяйке удовлетворенье.
Вот помидоры, чтоб крошить в салат,
Солить их в банках и варить томат.
Вот всей семьи кормилица – картошка,
Вот валерьяна для хозяйской кошки…
А прочим всем, кто пользы не несет,
Хозяйка тяпкой голову снесет.
Есть польза от тебя, скажи мне честно,
Даришь плоды иль украшаешь место?»
«Расту и всё!» «Так, братец, не пойдёт –
Не получает тот, кто не даёт!»
И люди есть, что сорнякам подобны –
Завистливы, жадны́, никчемны, злобны,
Всё думают, что им кругом должны,
А сами не милы и не нужны.
7. Полезные сорные растения. (рассказ)
Оказывается:
— сорняки очищают воздух;
— служат кормом для диких и домашних животных;
— являются пищевым продуктом для людей;
— полезны своими лекарственными свойствами;
— это сырьё для изготовления косметики;
— это плацдарм для появления на свет таких насекомых, как бабочки;
— это средство от вредных насекомых, и одновременно бытовой краситель и садовое удобрение;
— сорняки — отличные медоносы, они привлекают в сад насекомых, опыляющих «заодно» и садовые растения.
Сообщения учащихся о полезных свойствах сорняков.
II. Закрепление. Викторина по теме «Сорные растения». (Работа по группам).
Обведите кружочком правильный ответ. Каждое здание оценивается в 1 балл.
1. Как называется сорняк, изображенный на рисунке?
1) Лебеда
2) Полынь
3) Крапива
2. Где перечислены только сорняки?
1) Пырей, осот, крапива, астра, картофель.
2) Лебеда, мышиный горох, пырей, полынь.
3) Вьюнок, капуста, лебеда, подорожник.
3. Выберите верные высказывания.
1) Сорняки – это вредные бесполезные растения и их нужно уничтожать всеми способами.
2) Сорняки – это культурные растения, выращиваемые человеком.
3) Сорняки – это растения, которые приносят вред, засоряя сады, огороды, поля, но также имеют много полезных свойств и широко применяются в народной медицине.
4. Какие сорняки можно использовать в приготовлении салатов?
1) Одуванчик, крапива, щавель
2) Щавель, полынь, одуванчик.
5. Этот сорняк незаменим при царапинах, листик этого растения прикладывают к ранке для остановки крови и быстрого заживления.
1)Вьюнок полевой 2)Подорожник 3)Полынь
6. Этот сорняк лучше не брать голыми руками, можно обжечься.
1) Одуванчик 2) Щавель 3) Крапива
7. Отгадай по листику, что это за сорняк. Соедини стрелками название с листиком.
Вьюнок, одуванчик, подорожник, крапива, тысячелистник
8. Листья этого сорняка состоят из тысячи зубчатых пёрышек.
1) Тысячелистник 2) Крапива 3) Вьюнок
9. У этого растения стебель стелющийся, покрыт волосками, которые собирают в стебель влагу от полезных растений. Любит расти во влажных местах. Цветочки – мелкие белые.
1)Мокрица 2)Одуванчик 3)Крапива
10. Это самый обычный надоедливый сорняк. Засоряет все без исключения культуры, огороды, поля. Не зря говорят: «Осот да ………….. для посевов беда. В годы войны из этой травы пекли лепёшки, а из семян варили кашу.
1) Вьюнок 2) Мокрица 3) Лебеда
III. Итог занятия.
Не забывайте, что все в природе взаимосвязано. И если вы вдруг однажды удивитесь, почему это нынче так редки стали бабочки — знайте, что в значительной степени виной тому — ваше стремление вырастить сад без сорняков. На одном лишь из них на «отвратительной» вездесущей крапиве выкармливаются десятки видов этих прекрасных созданий: и дневной павлиний глаз, и бабочка-адмирал, и бабочка-медведица, и металловидка, и пестрокрыльница. Тщательно прополов свой сад, вы помогли цветам земли, но не оставили шансов для порхаюших «цветов».
Зимующие сорняки и меры борьбы с ними
Сорные растения – основные конкуренты сельскохозяйственных культур. Они засоряют поля и естественные кормовые угодья, а также делают невозможным существование культурных растений наравне с ними. Их вредоносность объясняется следующими факторами:
- они поглощают большое количество влаги и минеральных веществ из почвы, как следствие – снижают качество урожая;
- могут затенять культурные растения от солнца;
- многие сорняки являются источником возбудителей болезней, а также способствуют распространению насекомых-вредителей;
- усложняют уход за посевами, уборку урожая, засоряют комбайны.
Зимующие сорняки развиваются или как яровые, или как озимые растения. При прорастании семян весной они ведут себя как яровые сорняки, при появлении всходов летом и осенью — как озимые. Чаще всего зимующие сорняки засоряют озимые колосовые и многолетние травы. К представителям относят: пастушья сумка, ярутка полевая, василек синий, ромашка непахучая, крестовник обыкновенный, мак-самосейка и др.
Борьба с сорными растениями
Конечно, важнее всего не допустить развития сорняков на поле, проводить предупредительные меры борьбы: правильно чередовать культуры в севообороте, тщательно очищать посевной материал, соблюдать оптимальные нормы, сроки и способы посева, осуществлять своевременную и качественную уборку урожая и др. Но все же, если проблема засоренности имеет место, то для борьбы с сорными растениями используют механические, биологические и химические способы. Механический метод борьбы характеризуется применением специальных рабочих органов для культивации полей, их очистки от сорных растений. Помимо культивации проводится глубокая заделка их в почву при вспашке. В результате создаются такие условия, когда семена совсем не прорастают или их проростки гибнут, не достигнув поверхности, из-за истощения.
Биологический способ борьбы заключается в использовании насекомых, нематод, фитопатогенных микроорганизмов, которые поражают вегетативные и генеративные органы сорняков.
Химический способ борьбы подразумевает под собой использование гербицидов, которые будут подавлять развитие сорных растений активными действующими веществами. Это эффективный способ, который хорошо тем, что с его помощью можно работать с разной степенью засоренности и со всеми видами сорной растительности. Компания «Химагромаркетинг» для борьбы с зимующими сорняками предлагает применение сплошного гербицида Факел на основе глифосата кислоты (360 г/л). Он предназначен для борьбы с широким спектром сорняков на посевах. Первые его признаки действия можно наблюдать на 3-4 день: растения начинают желтеть и буреть. Полная гибель наблюдается на 7-14 день после обработки. Действующее вещество препарата попадает на зеленые части растений, проникает внутрь и тормозит его рост и развитие, а с течением времени растения и их корни погибают.
Преимущества данного препарата заключаются в том, что обработанные им растения не отрастают более, а также препарат не нарушает севооборот: после его использования можно высевать любые культуры. Кроме этого, Факел не имеет остаточного почвенного действия и не накапливается в объектах окружающей среды. В почве и воду под действием микроорганизмов активно распадается на простые соединения и, тем самым, является нетоксичным для млекопитающих, птиц, рыб, почвенных организмов и др.
Применение Факела минимизирует денежные и производственные расходы на обработку почвы, позволяет сократить их число, повысить влагообеспеченность и сэкономить финансовые и рабочие ресурсы. Препарат используют с нормой расхода от 4 до 8 л/га в зависимости от степени засоренности полей, и какие сорняки на них обитают.
Факел Экстра** — также продукт компании «Химагромаркетинг» — это гербицид сплошного действия на основе калийной соли глифосата в количестве 625 г/л. Факел Экстра** интересен тем, что характеризуется более высокой концентрацией действующего вещества – почти в 2 раза больше глифосата. А, значит, он работает быстрее, чем Факел, и интенсивнее уничтожает сорные растения на полях. Кроме этого, препарат адаптирован к более широкому температурному диапазону, и, таким образом, раздвигаются временные, биологически обоснованные рамки применения гербицида. Данный препарат можно применять с нормой расхода 1,0-4,0 л/га, при этом, опять же, нужно смотреть на засоренность полей и видовое разнообразие сорняков для определения оптимальной нормы расхода.
Гербициды сплошного действия просто необходимы для обеспечения чистого поля. Без их использования существует риск недружного прорастания всходов, а, значит, уже на данном этапе могут быть потенциальные потери в урожайности поля. Ведь известно, что именно в самом начале развития растений такой фактор, как превышающее количество сорняков на поле может существенно снизить продуктивность и качество сельскохозяйственных культур.
Гербициды же избирательного действия подавляют или уничтожают сорняки, не повреждая культурные растения. Их делят на почвенные и страховые (используемые по вегетации) гербициды.
Почвенные гербициды создают своеобразный «экран», который защищает поля от прорастания сорняков на срок 1-2 месяца в период всходов и начальные фазы развития, когда культурные растения не могут конкурировать за влагу, освещенность и питательные вещества в почве с сорняками.
Среди почвенных гербицидов стоит обратить внимание на такие препараты, как Тирон (метрибузин, 700 г/кг), который используется на томатах, картофеле и сое с нормой расхода от 0,5 до 1,4 кг/га. Хотя в общемировой практике применяется также на моркови, люцерне 2-го года и яблоне. Препарат борется с однолетними двудольными, в том числе с зимующими сорняками, а также злаковыми. Его механизм действия основан на ингибировании процесса фотосинтеза, а, значит, замедляется рост растений и вскоре они вовсе погибают.
Пульс-Стар** (имазамокс, 120 г/л) – почвенный гербицид, применяемый на сое, горохе, фасоли, а также подсолнечнике и рапсе, устойчивым к имидазолинонам. Действующее вещество проникает в сорные растения через корни и листья, накапливается и способствует снижению в растениях уровня аминокислот с последующим нарушением их синтеза. Рост сорняков с нормой расхода 0,25-0,4 л/га приостанавливается уже через несколько часов после обработки, а полная их гибель наступает через 3-6 недель после обработки, при этом действующее вещество Пульс-Стар** эффективно борется с зимующими сорняками.
Кроме этого, есть такой почвенный гербицид, как Позитив Плюс (прометрин, 500 г/л). Он борется с широким спектром сорных растений, в том числе с зимующими сорняками на подсолнечнике, кукурузе, картофеле, сое, горохе и зонтичных культурах. Прометрин блокирует процесс фотосинтеза в сорных растениях и при этом не является фитотоксичным для культурных растений при соблюдении нормы расхода (около 2,0-4,0 л/га). При этом препарат хорошо сочетается с почвенными противозлаковыми гербицидами.
Другой почвенный гербицид избирательного действия – Авангард (с-метолахлор, 960 г/л). Это системный препарат, который предотвращает прорастание сорняков и появление на поверхности почвы. Его действующее вещество вызывает нарушение естественных процессов в растениях, что приводит к его гибели. Авангард хорошо подходит для приготовления баковых смесей и характеризуется отсутствием фитотоксичности на широкий спектр культур. Рекомендуемая норма расхода 1,0-2,0 л/га, работает по таким культурам, как кукуруза, соя, подсолнечник, рапс, сахарная и столовая свекла, а также капуста.
Среди линейки послевсходовых гербицидов компании «Химагромаркетинг» следует обратить внимание на такие препараты, как:
Оптимум (дикамба кислота, 480 г/л) – применяется на зерновых культурах с нормой расхода 0,15-0,3 л/га, при этом он борется с зимующими сорняками, а также против однолетних двудольных сорняков, в том числе устойчивых к 2,4-Д и 2М-4Х. Визуальное действие гербицида проявляется через неделю-две после применения: листья скручиваются, увядают, а после все растение погибает. Данный препарат можно использовать на посевах кукурузы совместно с препаратом Приоритет – смесь эффективно работает против однодольных и двудольных сорных растений.
Приоритет (никосульфурон, 40 г/л), Сарацин (метсульфурон-метил, 600 г/кг) и Герсотил (трибенурон-метил, 750 г/кг) – системные гербициды, относящиеся к одному классу сульфонилмочевины. Они используются по вегетации и борются с зимующими сорняками. Приоритет работает на посевах кукурузы, а Сарацин и Герсотил – на посевах озимых и яровых колосовых культур. Эти гербициды угнетают рост сорных растений уже через 4-6 часов после обработки посевов. Действующее вещество поглощается всей поверхностью сорняков, при этом замедляются его процессы роста и развития, после чего растение погибает уже через 3 недели после обработки. Рекомендуемая норма расхода препарата никосульфурона – 1,0-1,5 л/га, трибенурон-метила – 0,015-0,025 кг/га, а метсульфурон-метила – 0,005-0,01 г/кг.
Цукрон + (клопиралид, 300 г/л) – также системный гербицид для защиты посевов от однолетних двудольных и многолетних корнеотпрысковых сорняков. Их рост останавливается через 2-3 часа после обработки. Первые признаки замедления роста визуально проявляются через 2-3 суток. Действие препарата основано на замедлении ростовых процессов, в результате чего растение погибает уже через 10-15 суток. Токсичный эффект достигается путем влияния препарата на процессы дыхания клеток. Рекомендуемая норма расхода препарата 0,1-0,66 л/га в зависимости от обрабатываемой культуры и степени засоренности.
2,4-Дактив (2-этилгексиловый эфир 2,4-Д, 860 г/л) – это также системный послевсходовый гербицид для борьбы с сорными растениями. Его скорость действия с нормой расхода 0,6-1,0 л/га составляет около 2-3 недель, причем первые признаки действия препарата заметны через 12-18 часов после опрыскивания, а окончательного уничтожения сорняков можно ожидать через 2-3 недели. Механизм его действия основан на нарушении процесса синтеза белков: препарат проникает в растение через надземные части и накапливается в нем, в результате чего нарушается водный обмен растения, теряется тургор, что приводит к гибели сорняка.
Химический метод борьбы — самый эффективный, который, однако, требует к себе ответственного подхода. Очень важно соблюдать регламентированные нормы применения препаратов, чтобы не только добиться максимального результата, но и не допустить появления резистентности у сорняков, а также загрязнения экосистем.
Конечно, для борьбы с зимующими сорняками лучше всего использовать не только химические, но и механические, и биологические методы борьбы, поскольку именно комплекс этих мероприятий будет способствовать наилучшему результату. Ну а если нужны именно химические препараты против сорняков, то компания «Химагромаркетинг» поможет Вам в их правильном подборе, проконсультирует Вас и с удовольствием ответит на все Ваши вопросы.
** — препарат на стадии регистрации, в портфолио компании войдет в 2019 году.
Размножение растений листовыми, тростниковыми и корневыми черенками
Некоторые, но не все, растения можно размножить с одного листа или с его части. Листовые черенки большинства растений не дают нового растения; они обычно дают только несколько корней или просто гниют. Поскольку листовые черенки не содержат пазушных почек, их можно использовать только для растений, которые способны образовывать придаточные почки.Листовые черенки используются почти исключительно для размножения некоторых комнатных растений. Выделяют несколько видов листовых черенков.
Удалите лист и включите до 1 1 ⁄ 2 дюймов черешка. Вставьте нижний конец черешка в среду (рис. 1). У основания черешка сформируется одно или несколько новых растений. Затем новые растения отделяют от исходного черенка листового черешка, и черенок можно использовать еще раз для получения большего количества растений.Примеры растений, которые можно размножать листовыми черенками, включают африканскую фиалку, пеперомию, эписцию, хойю и очиток.
Рисунок 1. Лист-черешок.
Этот метод используется для растений с толстыми мясистыми листьями.Змеиное растение (сансевиерия), однодольное, можно размножить, разрезав длинные листья на части размером от 3 до 4 дюймов. Вставьте черенки вертикально в среду. Африканскую фиалку, двудольную, также можно размножить с самой листовой пластинки. Срежьте с растения лист и удалите черешок. Вставьте лист вертикально в среду, убедившись, что средняя жилка погружена в среду для укоренения (рис. 2). Новые растения сформируются из средней жилки.
Рисунок 2.Лист без черешка.
Отделите лист от бегонии рекс и удалите черешок. Сделайте надрезы на нескольких выступающих жилках на нижней стороне листа (рис. 3). Положите черенок на материал нижней стороной вниз.Если лист скручивается, удерживайте его на месте, покрывая края средством для укоренения. На каждом срезе будут формироваться новые растения. Разновидностью этого метода является разрезание листа на клинья, чтобы на каждом кусочке была основная прожилка. Листовой клин следует вводить в среду с частичным прикрытием основной жилки.
Рисунок 3.Расщепленная жила.
Обрезки листовых почек используются для многих вьющихся лоз, а также при ограниченном пространстве или ограниченном материале для стрижки. Каждый узел на стебле можно рассматривать как черенок. Этот вид черенка состоит из листовой пластинки, черешка и короткого отрезка стебля с прикрепленной пазушной почкой.Поместите черенки в среду так, чтобы бутон был покрыт ( 1 ⁄ 2 на 1 дюйм) и лист был открыт (Рисунок 4). Примеры растений, которые можно размножать таким образом, включают клематис, рододендрон, камелию, нефритовое растение, каучуковое растение, дьявольский плющ, виноградный плющ, драцену, ежевику, махонию и филодендрон с сердцевинными листьями.
Рисунок 4.Листо-почковые черенки.
Черенки тростника — простой способ размножить некоторые заросшие, длинноногие комнатные растения, такие как тростник, кукуруза, китайское вечнозеленое растение и другие растения с толстыми стеблями.Секции стебля без листьев (от 2 до 3 дюймов в длину) вырезают из более старых стеблей. Каждая трость должна иметь один или два узла (рисунок 5). Положите черенок горизонтально на среду или вставьте его вертикально так, чтобы примерно половина черенка находилась ниже поверхности среды, оставив бутон вверх. Черенки тростника обычно пересаживают в горшок, когда появляются корни и новые побеги.
Рисунок 5.Обрезки тростника.
Некоторые растения можно размножать частями корня. Черенки древесных растений обычно берут с растений в период покоя, когда уровень углеводов высок.Корневые черенки некоторых видов дают новые побеги, которые затем образуют свою собственную корневую систему, тогда как корневые черенки других растений развивают корневую систему, прежде чем дают новые побеги. Примеры растений, которые можно размножить корневыми черенками, включают малину, ежевику, розу, лозу, флоксы, яблоню, инжир, сирень и сумах.
Растения с крупными корнями обычно выращивают в открытом грунте. Корневые черенки должны быть от 2 до 6 дюймов в длину. Сделайте прямой надрез на проксимальном конце (ближайшем к кроне родительского растения) и косой надрез на дистальном конце (самом дальнем от кроны) каждого корневого черенка.Свяжите черенки пучками все однотипными концами между собой. Важно соблюдать правильную полярность черенков. Хранить в течение 3 недель во влажных опилках, торфяном мхе или песке при температуре 40 ° F. Снять с хранения. Разместите черенки на расстоянии 2-3 дюйма друг от друга в хорошо подготовленной садовой земле. Верхушки черенков (проксимальные концы) должны быть на 2-3 дюйма ниже поверхности почвы.
Для растений с небольшими корнями разрежьте корни на секции от 1 до 2 дюймов. Положите черенки горизонтально на среднюю поверхность на ровной поверхности и засыпьте примерно 1 ⁄ 2 дюймов почвы или песка.Поместите квартиру в полиэтиленовый пакет или накройте стеклом. Поместите квартиру в тень и снимите защитный кожух после появления новых всходов.
- Bryant, G. 1995. Справочник распространения . Книги Стэкпола: Механиксберг, Пенсильвания.
- Дирр, М. А. и К. В. Хойзер, мл. 1987. Справочное руководство по размножению древесных растений: от семян к культуре тканей .Университетская пресса: Афины, Грузия.
- Хартманн, Х. Т., Д. Э. Кестер, Ф. Т. Дэвис и Р. Л. Женева. 1996. Размножение растений, принципы и практика . 6-е изд. Прентис Холл: Верхняя Сэдл-Ривер, Нью-Джерси.
- Макмиллан Обзор, П. Д. А. 1978. Размножение растений . Саймон и Шустер: Нью-Йорк.
- Toogood, A. 1993. Размножение растений стало проще . Timber Press: Портленд, Орегон.
- Эрвин Эванс
- Консультант по вопросам садоводства (потребительское садоводство)
Наука о садоводстве
- Фрэнк Блазич
- Профессор
Садоводство
Дополнительную информацию можно найти на следующих веб-сайтах NC State Extension:
Дата публикации: янв.31, 1999
N.C. Cooperative Extension запрещает дискриминацию и домогательства независимо от возраста, цвета кожи, инвалидности, семейного и семейного положения, гендерной идентичности, национального происхождения, политических убеждений, расы, религии, пола (включая беременность), сексуальной ориентации и статуса ветерана.
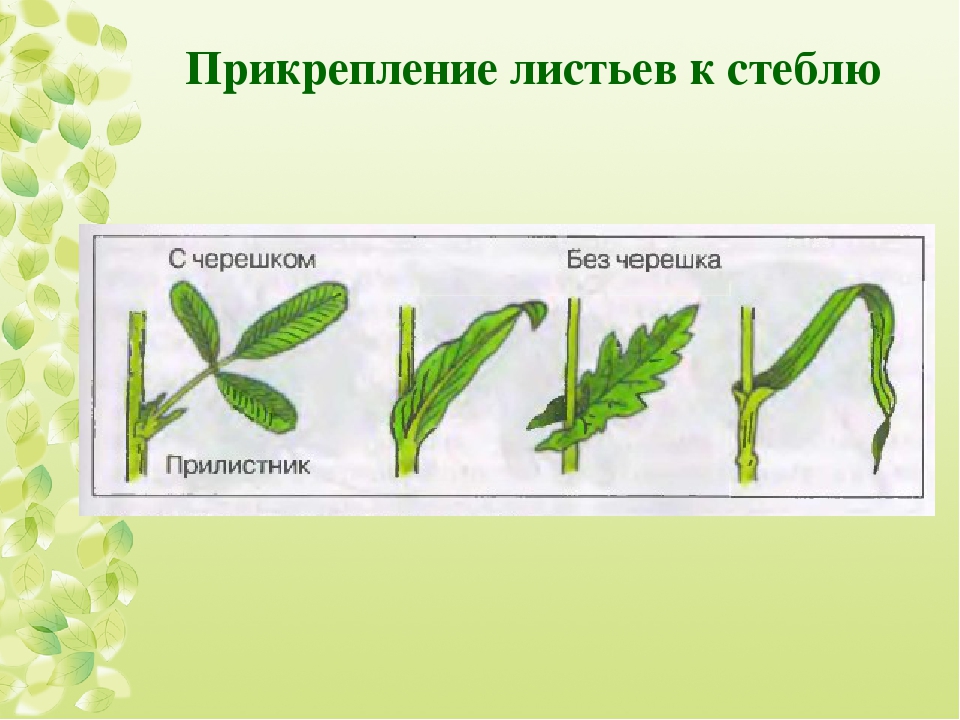
Идентификация растений: изучение листьев | Пейзажные растения
Чтобы идентифицировать элемент, необходимо распознать элемент и связать его с соответствующим именем. Например, этот коричневый автомобиль перед нашим домом — это Хонда Аккорд. Или это большое древесное растение в парке — это дерево, а точнее пихта Дуга. Для идентификации ландшафтного или садового растения необходимо распознать растение по одной или нескольким характеристикам, таким как размер, форма, форма листьев, цвет цветка, запах и т. Д., И связать это распознавание с именем, либо общим, либо так называемым научным названием. . Точная идентификация культурного растения может быть очень полезной для понимания того, как оно растет (например,г., размер, форма, текстура и т. д.), а также способы ухода и защиты от вредителей и болезней.
Сначала давайте рассмотрим около общих характеристик растений, которые можно использовать для их идентификации . Если бы это был класс ботаники , занимающийся систематикой растений , областью исследований, связанной с идентификацией, наименованием, классификацией и эволюцией растений, мы бы потратили много времени на репродуктивных частей растений , т. Е. в основном различные части цветка , т.е.е., тычинки, завязь, рыльце и т. д. Структурное сходство репродуктивных частей является важным средством, с помощью которого растения классифицируются, группируются, называются и, следовательно, идентифицируются. Однако для многих садовых растений, особенно для древесных растений , нам, возможно, придется идентифицировать без учета цветов , поскольку часто цветы отсутствуют или они очень маленькие, а другие характеристики могут быть более очевидными. Некоторые характеристики растений настолько очевидны или уникальны, что мы можем распознать их без детального изучения растения.Точно так же, наверное, все мы сразу узнаем Volkswagen Beetle среди группы автомобилей на стоянке.
Итак, какие характеристики растений можно использовать для идентификации растений?Листья часто являются основой для идентификации растений. , так как их легко увидеть. Обычно они состоят из двух частей ,
- лезвие , широкая или более заметная часть полотна,
- и «стебель» или черешок , с помощью которого лезвие прикреплено к стеблю.Есть бутон в точке , где черешок прикрепляется к стеблю [см. Ольха Ситкинская, Alnus viridis subsp. sinuata , лист и почки]. (Чтобы вернуться на эту страницу, воспользуйтесь кнопкой Назад в своем браузере.)
Во-первых, имейте в виду, что все листья на данном растении не имеют одинаковый размер или даже внешний вид [ Betula , сравнение размеров листьев]. Они могут различаться по размеру, цвету и даже форме [ Sassafras albidum , листья, осень] и [ Malus sargentii , листья, осень]; Те, кто получает много солнца, могут отличаться от тех, кто находится в густой тени.Поэтому, пытаясь определить идентичность растения по его листьям, убедитесь, что вы исследуете много листьев и попытаетесь определить, что можно считать «типичными» характеристиками листа. Хотя баскетболисты могут различаться по размеру, форме и цвету, «типичной» физической характеристикой баскетболиста часто является «рост».
Широкие и узкие створки
Листья можно разделить на широкие и узкие.
- Широкие листья имеют широкую пластинку , часто с видимой сетью прожилок.
Северная Катальпа, [ Catalpa speciosa , листья и цветы].
Знакомые примеры растений с широкими листьями: яблоня ( Malus ), дуб ( Quercus ), клен ( Acer ) и т. Д.
- Узкие листья — это тонкие, без широкого лезвия , эти листья часто называют «иголки» или «чешуйчатые» .
Хвойные деревья, такие как сосна ( Pinus ), ель ( Picea ) и можжевельник ( Juniperus ), имеют узкие листья, некоторые имеют иголок
Ель обыкновенная, [ Picea abies , ветвь, хвоя]
Кедр Деодар и Кедр Голубой Атлас, [ Cedrus deodara и Cedrus atlantica ‘Glauca’, сравнение ветвей]
и другие имеют чешуйчатых листьев,
Кедр благовонный, [ Calocedrus decurrens , веточки с чешуевидными листьями,
Западный красный кедр, [ Thuja plicata , веточки с чешуевидными листьями].
Приставной лист
Шаблон , по которому листья прикрепляются к стеблю или ветке , также является полезной характеристикой при идентификации растений. Есть две большие группы: , альтернативные и , противоположные образцам , и третья, менее распространенная модель, с завитками .
- Альтернативные листья имеют только один лист , прикрепленный в одном месте (узел) на стебле, часто листья чередуются с одной стороны на другую при движении вдоль стебля, или они могут иметь спиральный узор вокруг стебля. корень.
Красный бутон восточный, [ Cercis canadensis , листья]
Вяз американский, [ Ulmus americana , листовой побег]
Чередующиеся листья распространены у следующих родов: Alnus (ольха), Crataegus (боярышник), Cotoneaster , Magnolia , Prunus , Quercus (дуб) и Rubus .
- Листья, расположенные напротив , относятся к , два листа прикреплены в одном и том же месте (узел) на стебле, , но напротив друг друга , то есть по обе стороны от стебля
Самшит обыкновенный, [ Buxus sempervirens , листовые побеги]
Katsuratree, [ Cercidiphyllum japonicum , листья]
Dawn Redwood, [ Metasequoia glyptostroboides , хвоя, сравнение].
Противоположные листья обычны у следующих родов: самшит (самшит), Cornus (кизил), бересклет , Fraxinus (ясень), Lonicera и Viburnum. Все клены ( Acer ) имеют супротивные листья.
- Иногда более двух листьев возникают из одного и того же места (узел) на ветке, листья могут исходить из ветки, как спицы на колесе, это называется расположением с завитками .
Redvein Enkianthus, [ Enkianthus campanulatus , побеги].
- Иногда данное растение может демонстрировать более одного типа расположения листьев. Например, у Crape Myrtle ( Lagerstroemia indica ) нижние листья побега могут иметь противоположное расположение листьев, но ближе к концу побега листья могут быть чередующимися или даже мутовчатыми [ Lagerstroemia indica , побег].
Листы простые и сложные
Листья могут иметь одну нераздельную пластинку или пластину, разделенную на части.
- Простые листья имеют только одну листовую пластинку, со стеблем или черешком или без них.
Ольха белая, [ Alnus rhombifolia , листовой побег]
Клен красный, [ Acer rubrum , побег, листья]
Мирт обыкновенный, [ Myrtus communus , лист]
- Сложные листья имеют более одной пластинки и могут иметь сложную структуру стебля листа.
Paperbark Maple, [ Acer griseum , листья]
Медокуст без шипов, [ Gleditsia triacanthos var.инермис , л.].
Существует несколько различных типов сложных листьев, самые распространенные из них:
- Пальчатые сложные листья имеют три или более листочка, прикрепленных к концу стебля (черешка) (как пальцы на наших руках).
Огайо Бакай, [ Aesculus glabra , лист]
Целомудренное дерево, [ Vitex agnus-castus , лист].
- Перистые сложные листья имеют несколько листочков, прикрепленных вдоль центрального стебля.
Желтое дерево американское, [ Cladrastis kentukea , лист]
Небесное дерево, [ Alilanthus altissimus , лист].
Также они могут быть:
- двойное перистое ( двупородное ) соединение,
Шелковое дерево, [ Albizia julibrissin , лист]
Kentucky Coffeetree, [ Gymnocladus dioicus , лист] или
- тройное перистое (, трипинатное, ) соединение.
Небесный бамбук, [ Nandina domestica , лист].
Помните,
- листьев имеют бутон у основания стебля (черешка) , например, Paperbark Birch, [ Betula papyrifera , побег, листья], тогда как
- листовок нет, например, American Yellowwood, [ Cladrastis kentukea , leaf].
не всегда легко найти почку у основания черешка, она может быть не видна на ранней стадии роста, а иногда зрелая почка «спрятана», например, окружена основанием черешка, например, у
Ароматный снежный колокольчик, [ Styrax obassia , черешок листа и закрытая почка]
London Planetree, [ Platanus × acerifolia , черешок и бутон],
American Yellowwood, [ Cladrastis kentukea , стебель и бутон листа, осень].
Посмотрите на весь побег, чтобы определить, что это за лист, а не просто смотрите на конец ветки. Поскольку почка находится у основания каждого листа, можно определить расположение листьев (т. Е. Чередование, противоположность и т. Д.) Листопадного растения зимой, посмотрев расположение бутонов на голой ветке, например, красной Клен, низкорослые растения противоположного расположения, [ Acer rubrum , побеги ветвей и почек, зима].
Доли листа
Листья могут быть лопастными или недолстыми. Лепесток может быть определен как изогнутый или закругленный выступ . У листьев нет четкого различия между мелкими лопастями и глубокими зубцами. Основная вена часто видна в мочке, а в зубах этого может не быть.
Листья лопастные:
Живая изгородь, [ Acer campestre , листья и плоды]
Клен амурский, [ Acer ginnala , листья и плоды]
Клен крупнолистный, [ Acer macrophyllum , лист, осень]
Дуб Орегонский белый, [ Quercus garryana , лист]
Листья без лепестков:
Pacific Serviceberry, [ Amelanchier alnifolia , листья, осень]
Western Catalpa, [ Catalpa speciosa , листья, осень]
Eastern Redbud, [ Cercis canadensis , лист].
Листовое поле
Другой важной характеристикой листа для идентификации растения является край или край листа или листочка. Листья имеют либо гладкие края, называемые целиком , либо небольших выемок или «зубцов» по краю.
Форзиция белая, [ Abeliophyllum distichum , листья]
Дерево Небес, [ Ailanthus altissima , листочки] (в основном цельные, но с несколькими грубыми зубцами).
- Зубчатый : Зубцы могут находиться у основания листа, на кончике или по всему краю. Зубы могут различаться по количеству и размеру.
- Зубчатый , грубый, может быть трудно отличить от лопастного , например, Paperbark Maple, [ Acer griseum , листья и плоды]
- Зубчатый, дважды , Ольха ситкинская, [ Alnus viridis subsp. sinuata , край листа, поверхность]
- Зубчатая передача: зубчатая пила , зубья направлены вперед
- Однопильчатая ;
Zelkova японская, [ Zelkova serrata , листовые побеги]
Земляничное дерево, [ Arbutus unedo , листья] - Двузубчатый , Вяз американский, [ Ulmus americana , край листа и кончик]
- Колючий пильчатый , Барбарис Винтергрин, [ Berberis julianae , лист]
- Однопильчатая ;
- Зубчатые: с краевыми зубами, вершины которых перпендикулярны краю и не обращены вперед, Crimson Glory Vine, [ Vitis coignetiae , лист]
Другие характеристики листа, которые следует учитывать, особенно при использовании ботанического ключа.
- по всей форме (например, эллиптическая, ланцетная, линейная, обратнояйцевидная, продолговатая и т. д.)
- форма основания (клиновидная [клиновидная], сердцевидная, округлая и др.)
- форма вершины (крутая, заостренная, острая, выемчатая, мукронатная и т. д.)
- рисунок прожилок (например, параллельный, сетчатый и т. д.)
- свойства поверхности (например, опушенная, гладкая [гладкая])
- запах при раздавливании (сильный, неприятный, отсутствует и т. д.)
Нелистовые характеристики также полезны при попытке идентифицировать древесные растения, к ним относятся:
- тип цветка, цвет и эффектность
- тип, форма и цвет плодов при созревании.
В виде масштаба
- Чешуйчатые листья обычно мелкие, короткие, перекрываются ; они обычны для нескольких родов хвойных деревьев, включая можжевельники ( Juniperus ), falscypress ( Chamaecyparis ) и туи ( Thuja ), например, Arborvitae и Western Red Cedar ( T.plicata ), [ Thuja , веточки, сравнение]. Часто чешуйчатые листья отображаются по два, три или четыре на узел . Для этого часто требуется ручной объектив или маломощный микроскоп.
Различия в чешуйчатых листьях можно использовать для различения следующих «кедров», произрастающих в Орегоне (ни один из которых не является настоящими кедрами, т. Е. Cedrus ).
- Кедр благовоний [ Calocedrus decurrens , листья].Обратите внимание, что 4 листа появляются на той же ноте , 2 лицевых (лицевых) и 2 боковых (боковых), контур пары боковых листьев образует «дымовой бокал для вина».
- Порт-Орфорд Кедр [ Chamaecyparis lawsoniana , листья]. Листья плотно прижаты друг к другу супротивными парами . Боковые листья крупнее лицевых, там, где они встречаются на нижней стороне веточки, видна белая восковая линия, обозначенная знаком «X». Также обратите внимание на одну точку, смоляную железу, заметную на каждом листе лица (для этого может потребоваться ручная линза).
- Кедр красный западный [ Thuja plicata , листья]. Примечание: 4 листа одинакового размера (2 лицевых и 2 боковых) появляются на узле, восковые поверхностные отметины на нижней стороне веточки, как полагают, напоминают «бабочку» или «галстук-бабочку».
- Кедр желтый или аляскинский [ Chamaecyparis nootkatensis , листья]. Листья плотно прижаты друг к другу в противоположных парах , (лицевая пара и боковая пара), 2 листа в данном узле имеют одинаковый размер, на стыке листьев не наблюдается восковой маркировки.
Вот сравнение четырех «кедров», произрастающих в Орегоне.
Игольчатые листья, также распространены у хвойных пород, прикрепляются к веткам несколькими способами:- Одноместный —
- крепится прямо к ветке , например,
- прикреплен с помощью стержневой ножки , например,
- Связки — , сгруппированные в связки , прикрепленные к ветке; часто бывает 2, 3 или 5 листьев в пучке.У данного дерева обычно одинаковое количество иголок в пучке. Пучки обычны у сосны:
2 иглы / пучок, австрийская сосна, [ Pinus nigra , хвоя и зрелые шишки]
3 иглы / пучок, сосна Ponderosa, [ Pinus ponderosa , иглы и зрелая шишка]
5 игл / пучок, западная белая сосна, [ Pinus monticola , хвоя]
- Кластеры — обычно более 5, может быть 30 и более , например
Кедр Деодар ( Cedrus deodara ) [ Cedrus deodara , пучок игл] и
Кедр Голубой Атлас ( Cedrus atlantica ‘Glauca’), [ Cedrus atlantica ‘Glauca’ , пучок игл].
Примечание:
Пихта плоские иглы приятны на ощупь , обычно, но [испанская пихта остро острая]
Ель острая, квадратная (иглы в поперечном сечении)
Сосна в упаковках (иглы группами по 2, 3, 5, реже по одной)
Ссылки :
Cope, E.A. Ключ Мюншера к древесным растениям: расширенное руководство по местным и культивируемым видам.2001. Издательство Корнельского университета, Итака, Нью-Йорк.
Partyka, R.E., J.W. Римельспах, Б. Джойнер и С.А.Карвер. 1980. Древесные орнаментальные растения: растения и проблемы. ChemLawn Corp. Колумбус, Огайо.
Вьющиеся растения с усиком на черешке (усики на черешке)
Опоры для лазания
За некоторыми исключениями (C lematis vitalba и Clematis montana) у черешковых вьющихся растений не развивается прочная структура стебля. По этой причине для них требуются легкие и хрупкие опорные системы с довольно мелкой сеткой (небольшой шаг сетки).Оптимальным вариантом являются стальные канаты, но также очень подходят тонкие стержни, которые используются для бамбуковых шпал . Более толстые стержни или посохи, такие как те, что используются в деревянных шпалерах, требуют использования растительных стяжек . Подходят самые тонкие сетки — решетчатая сетка (сделанная из синтетических материалов, таких как пластик или нейлон) или проволочная сетка (например, проволочная сетка), но недостатком является то, что кусочки одревесневших усиков и побегов накапливаются каждый год и, как правило, оседают. застревает в сетке, что делает удаление немного более утомительным.
Подходящие системы троса
Системы троса с размером ячеек прибл. 25-30 см, как и канатные системы 5040 или 5050 , являются наиболее подходящими. Это нормально, если преобладают вертикальные веревки, так что сетки образуют вертикальные прямоугольники, а не квадраты. Обычно достаточно средних и канатных систем или даже easy и light .
Тросовые системы с ограниченной пригодностью
Одиночные вертикальные тросы не очень подходят, поскольку они не обеспечивают достаточного сцепления с оборудованием.Точно так же конструкции с одной горизонтальной проволокой наверху, где цветы должны без ограничений занимать все необходимое им пространство, не очень подходят. Также ограниченно пригодны: системы с преимущественно горизонтальными проволоками и слишком широкими размерами ячеек; с последним становится невозможным охватить территорию целиком. Однако, если вы хотите добиться определенного эффекта роста, например, нависания / каскадного роста , вы можете сделать это с помощью этих типов кабельных систем.
Системы с очень маленькими размерами ячеек или плотно расположенными проводами также не являются оптимальными, поскольку они не только дороги, но и не дают предприятию дополнительных преимуществ. Требуемое обслуживание (удаление усиков) было бы дополнительным недостатком.
Неподходящие канатные системы
Канатные системы, которые недостаточно велики / высоки, чтобы соответствовать ожидаемой высоте роста соответствующих видов растений, также явно не подходят.
Листовая зелень и черешки — растительные ресурсы Растительные ресурсы
Листовые овощи (группа культур 4) включают как листовую зелень (например, шпинат), так и черешки (сельдерей) для свежего сбыта и переработки.Звездочки указывают на репрезентативные культуры Группы.
Листовая зелень
Одуванчик . Произведен в LRGV для придорожной и коммерческой продажи. Розетки культурных видов могут иметь ширину от 18 до 24 дюймов. Собирают вручную через два-три месяца и связывают пучками. Используется как зелень и в салатах; сушеные корни (Taraxacum) в лечебных целях. Несколько листовых насекомых и патогенов, в том числе мучнистая роса.
Эндивий и цикорий . Прохладный однолетник с распущенными колосьями.У эндивия продолговатые бахромчатые листья. Кудрявые виды — это цикорий. Широкие плоские листья — это эскарол. Выращивается так же, как шпинат, обычно на небольших посадках для продажи на обочине дороги или на местном рынке для местного использования в салатах или травниками. Родом из Средиземноморья. Некоторые кормушки для листовых насекомых. К болезням-вредителям относятся антракноз, мозаика, корневая гниль хлопчатника, ржавчина, гниль листьев и конечностей.
Салат * . Самый популярный из всех листовых овощей. Около 350 акров всех типов; кочанный салат (айсберг) в LRGV и High Plains; некоторые виды листовых или роменских, зеленолистных, краснолистных и других новинок в Зимнем саду.Качество и внешний вид имеют первостепенное значение. К насекомым-вредителям относятся тля, минеры и клещи. К сорнякам относятся озимые однолетники. Болезни включают Botrytis, гниение, ложную мучнистую росу, комплекс вирусов мозаики и мягкую гниль.
Петрушка свежая . Холодостойкий двулетник, убранный в первый сезон после посадки. Выращивается в районе LRGV для продажи на свежем рынке. Вредители похожи на редис и репу. Сушеная петрушка входит в состав лечебных трав (Группа 19).
Шпинат * .Техас занимает второе место по производству в США. Когда-то выращивался на 40 000 акров, теперь от 3 000 до 7 000 акров на площади WG . Сажают с сентября по ноябрь для многократного сбора урожая с ноября по апрель. На территории Зимнего сада; 70% — это гладколистные сорта для переработки. В LRGV и High Plains 20% составляют савойские или морщинистые листья. Устойчивость к болезням благодаря селекционным программам в Арканзасе и семеноводческим компаниям. Вредители приводят к серьезным потерям урожайности и качества. Качество чрезвычайно важно, поскольку в консервированных продуктах не допускаются черви и тля, а в собранном урожае не должно быть сорных частей растений.К сорнякам относятся поросенок, портулака, однолетние травы, зимние однолетние широколиственные, осот и мальва. К насекомым-вредителям относятся свекла и совка, проволочники, белые личинки, личинка кроны, петрушка для капусты, клещи, тля, огненные муравьи (уничтожающие семена на посадках) и огуречный жук. Болезни включают белую ржавчину, ложную мучнистую росу, пятнистость листьев альтернариоза, отшелушивание, антракноз, курчавую верхушку, вирусы западной желтизны свеклы и угасание фузариоза. Потрясающий профиль урожая «говорит все» о вредителях шпината и защите.
Черешок овощной
Сельдерей * . Выращено на LRGV для продажи в свежем виде. Желательны жесткие, компактные, прямые стебли с ярким блеском. К насекомым-вредителям относятся тля, минеры и клещи. К сорнякам относятся ореховая ослица и однолетние сорняки. Заболевания включают бактериальный ожог, пятнистость листьев, раннюю и фитофтороз, отмирание, ризоктонию и склеротинию.
Кинза . Кинза включена в раздел Травы (Группа культур 19) вместе с кориандром.
Швейцарский мангольд . Свекловидная культура, прямой посев, выращивание на листовых пластинах и
черешках. Зеленые сорта составляют 78% продукции; 22% — красные типы. Вредители, аналогичные вредителям свеклы и других зеленых культур.
| Урожай | Производство в масштабе штата | Площадь по регионам производства | ||||||
|---|---|---|---|---|---|---|---|---|
| акров | долларовая стоимость за акр | Общая стоимость (в тысячах долларов) | Нижняя долина | Зимний сад | Область равнин | Фар-Уэст, Техас | Восточные районы | |
| Зелень | ||||||||
| Одуванчик | 200 | $ 2 800 | $ 560 | 200 | 0 | 0 | 0 | 0 |
| Эндив | 150 | $ 1,600 | 240 | 150 | 0 | 0 | 0 | 0 |
| Салат * | 350 | 2200 долларов США | $ 770 | 100 | 150 | 50 | 0 | 50 |
| Петрушка | 700 | $ 4 200 | 2 940 долл. США | 700 | 0 | 0 | 0 | 0 |
| Шпинат * | 4 420 | 3 900 долл. США | $ 17 238 | 300 | 3,500 | 500 | 0 | 120 |
| Черешки | ||||||||
| сельдерей * | 200 | $ 9 700 | 1 940 долл. США | 200 | 0 | 0 | 0 | 0 |
| Швейцарский мангольд | 300 | 3 200 долл. США | $ 960 | 250 | 50 | 0 | 0 | 0 |
| Итого | 6,320 | 3 900 долл. США | 24 648 долл. США | 1 900 | 3,700 | 550 | 0 | 170 |
Анализ растений для сельскохозяйственных культур
Отбор образцов растений в начале сезона
Анализ только концентрации питательных веществ обычно неэффективен при диагностике многих проблем.Расчет усвоения питательных веществ — лучший выбор. Почему? Питательные вещества, даже если один или несколько может быть недостаточным, обычно больше концентрируются в низкорослых растениях. Например, концентрация азота может быть выше у растений высотой 12 дюймов по сравнению с гораздо более высокими растениями. В более высоких растениях азот просто разбавляется углеводами. Расчет усвоения питательных веществ — лучший подход. Чтобы рассчитать потребление питательных веществ, убедитесь, что вы:
1) высушить все собранные растения,
2) получите точный вес, а
3) завершите анализ растительного материала.
Чтобы рассчитать потребление питательных веществ, умножьте сухой вес растения на концентрацию питательных веществ. Зная количество отобранных растений, можно определить поглощение для отдельного растения. Для измерения поглощения питательных веществ должен быть доступ к печи, которая будет быстро сушить образец, и к весам или электронным весам, которые могут измерять небольшие различия в весе. Итак, необходимо некоторое планирование, если есть намерение рассчитать потребление питательных веществ. В диагностических ситуациях собирайте пробы почвы всякий раз и везде, где вы собираете пробы растений.Анализ образцов почвы часто может служить хорошим индикатором дефицита питательных веществ. Сравнивая результаты анализа собранных образцов почвы, вы можете подтвердить или отклонить предполагаемый дефицит питательных веществ.
Отбор проб растений в конце сезона
При отборе проб в конце сезона меньше внимания уделяется общему усвоению питательных веществ и больше внимания уделяется отбору проб частей растений, которые при сравнении концентрации питательных веществ хорошо коррелируют с конечной урожайностью сельскохозяйственных культур.Для кукурузы: образцы листьев расположены напротив и ниже колоса при появлении шелка при опадании пыльцы. Выбор времени для сбора образцов кукурузы очень важен. Соберите образцы до того, как шелк станет коричневым. Концентрация питательных веществ существенно снижается после этого момента жизненного цикла, и признанные стандарты не могут использоваться для сравнения. Для соевых бобов стандартом является образец трифолиатов, созревших недавно, от начала до середины цветения.
Отбор проб растений как средство диагностики
При использовании в качестве диагностического инструмента мы ожидаем, что анализ растений позволит выявить предполагаемый дефицит питательных веществ.В таких ситуациях мы обычно сталкиваемся с нормальными и низкорослыми и / или неокрашенными растениями на одном поле. Обычно люди собирают низкорослые растения и проводят анализ растительной ткани. Однако отбор образцов растений является более сложным, если мы ожидаем, что анализ тканей будет эффективным диагностическим инструментом. Для успешной идентификации возьмите три образца. Соберите один образец целых растений из низкорослого участка. Затем возьмите образец целых растений из краевой области, где наблюдается небольшое снижение роста или где растения немного отстают в росте.В третьем примере используйте нормальные и здоровые растения.
Отбор проб растений как инструмент мониторинга кукурузы и сои
Анализ растений также можно использовать для оценки уровня питательных веществ растений в зависимости от используемой программы удобрений. При использовании для этой цели методы взятия проб отличаются. Это обсуждение будет сосредоточено на кукурузе и сое. Поскольку результаты анализа растений будут сравниваться с известными стандартами, части растений следует отбирать на определенном этапе развития.Результаты анализа этих образцов ткани сравниваются со стандартами, которые приведены в таблицах 2 и 3.
Анализ растений при правильном использовании может стать полезным инструментом управления в современном сельском хозяйстве. Чтобы получить достоверную информацию, остановитесь и подумайте, прежде чем собирать образцы. Вы собираете образцы для диагностики проблемы или отслеживания результатов программы удобрений?
Границы | Реакция растений на близость к растительности: вся жизнь избегает тени
Введение
За последние несколько десятилетий повышение урожайности произошло в основном за счет увеличения плотности посадки.При текущем сценарии быстрого роста населения и ограниченного количества пахотных земель на Земле (http://www.fao.org/news/story/en/item/35571/icode/) эффективные методы ведения сельского хозяйства могут потребовать еще более высокой плотности посадки. вместе с изменениями в архитектуре растений как способ максимизировать урожайность. Оптимальная плотность посадки каждого вида растений определяется специфическим набором требований и факторами окружающей среды, которые напрямую влияют на рост растений и, следовательно, урожайность сельскохозяйственных культур.Однако в целом по мере увеличения плотности посадки растут и тени, подаваемые растениями, и, следовательно, активация реакции растений на близость растительности. Для многих видов растений восприятие близости растений приводит к набору реакций, известных как SAS, которые сильно влияют на развитие и архитектуру растений. При активации SAS приводит к уменьшению завязывания семян и изменению развития плодов. Зерновые культуры обычно выращивают при высокой плотности, и ответы SAS, если они активированы, обычно вредны для урожайности, потому что они вызывают перераспределение ресурсов в сторону удлиненного роста за счет собираемых органов (обычно листьев, плодов и семян).Следовательно, одна из ключевых исследовательских задач, которые, безусловно, окажут большое влияние на сельское хозяйство, — это выявление механизмов, лежащих в основе развития растений в ответ на близость растительности.
Цель этого обзора — описать то, что известно о реакции на тень во взрослых тканях, таких как черешки взрослых листьев, а также немного информации о реакции SAS в репродуктивных тканях. Мы сосредоточены на исследованиях, проведенных на Arabidopsis thaliana в ответ на передачу сигналов красным (R, около 600-700 нм) и FR (около 700-800 нм), как влияние PAR (между 400 и 700 нм) и других факторов окружающей среды. факторы, такие как зеленый и синий свет, производство этилена или механические стимулы, были недавно рассмотрены (Pierik and de Wit, 2014).Мы обсуждаем ответы, происходящие во взрослых тканях, на основе имеющихся знаний о молекулярных механизмах и компонентах, играющих роль в регуляции SAS-ответа гипокотилей и их участии в транскрипционных сетях, которые опосредуют этот процесс.
Сигналы приближения к растительности и восприятие растений
Как в естественных, так и в сельскохозяйственных растительных сообществах свет может стать ограничивающим ресурсом при высокой плотности растений. В такой ситуации растения эволюционировали, чтобы либо терпеть, либо избегать тени.Теневыносливость достигается за счет различных наборов реакций у разных видов, таких как изменения в физиологии листьев, биохимии, анатомии и морфологии и / или архитектуре растений. В целом, при низкой интенсивности света теневыносливые виды склонны адаптироваться к очень консервативному использованию ресурсов, что обычно сопровождается низкой скоростью роста, более тонкими листьями, сниженным апикальным доминированием (повышенным ветвлением) и низкой реакцией удлинения (Martinez-Garcia et al. , 2010; Casal, 2012; Gommers et al., 2013). Напротив, виды, избегающие тени (или любящие солнце), обычно склонны адаптировать свое развитие, чтобы способствовать удлинению за счет развития листьев и увеличивать верхушечное доминирование (уменьшение ветвления), позволяя молодым и растущим тканям ускользать из тени. Эти реакции SAS можно рассматривать как оптимальные стратегии адаптации к возможному затенению в естественной среде. Напротив, активация SAS обычно пагубно сказывается на урожайности сельскохозяйственных культур с агрономической точки зрения.
Как растительность заявляет о себе: сигнал R: FR
В открытых условиях, т.е.е., когда растение растет в условиях низкой плотности растительности, свет, исходящий от солнца (или солнечного света), имеет относительно постоянное качество, а R: FR составляет примерно 1,2–1,5. В среде с высокой плотностью растительности могут возникать две связанные, но разные ситуации, которые снижают R: FR: близость к растению (без прямого затенения соседними растениями) и прямая тень растительного покрова . Поскольку считается, что растительность предпочтительно отражает FR по сравнению с другими длинами волн, близость растений вызывает промежуточное или умеренное снижение R: FR.Напротив, под пологом растений фотосинтетические пигменты из зеленых тканей специфически поглощают свет из области PAR (между 400 и 700 нм), тогда как FR, который плохо поглощается тканями растений, отражается от растений или проходит через них. Следовательно, под прямым затенением растительного покрова как количество PAR, так и R: FR сильно снижается (низкий и очень низкий R: FR), последний эффект в основном связан с избирательным истощением R солнечного света, фильтруемого листьями (Смит , 1982; Мартинес-Гарсия и др., 2010; Casal, 2013).
Как растения воспринимают близлежащую растительность: роль фитохромов
Изменения R: FR, указывающие на близость растения или оттенок, обнаруживаются фоторецепторами фитохромов, которые в A. thaliana кодируются небольшим семейством генов из пяти членов (от PHYA до PHYE ). В то время как PHYA кодирует единственный фотолабильный фитохром, phyA, остальные гены PHY кодируют фотостабильные фитохромы, phyB-phyE. Фитохромы существуют в двух фотопреобразованных формах: в неактивной форме Pr, поглощающей R, и в активной форме Pfr, поглощающей FR.Следовательно, на свету установившееся соотношение форм Pr и Pfr (которое также можно описать как относительные уровни активной формы Pfr, Pfr / Ptot) зависит от R: FR. Действительно, существует положительная корреляция между уровнями R: FR и Pfr / Ptot. При высоком R: FR (то есть при низкой плотности растительности) фоторавновесие смещается в сторону активной формы Pfr, Pfr / Ptot высокое (0,7 или выше), и SAS подавляется. Напротив, при низком R: FR фоторавновесие смещается в сторону неактивной формы Pr, Pfr / Ptot низкое (0.6 или ниже), а САС индуцируется (Smith, 1982, 2000; Casal, 2012, 2013; Possart et al., 2014).
Генетический анализ Arabidopsis показал, что мутанты, дефицитные по phyB, демонстрируют длинные гипокотили и раннее цветение при выращивании в условиях высокого R: FR (в открытых условиях или при низкой плотности растительности). Эти фенотипы очень похожи на фенотипы растений дикого типа, растущих при низком R: FR (рис. 1). Следовательно, конститутивный фенотип SAS мутантных растений phyB был принят как показатель того, что phyB репрессирует ответы SAS в условиях высокого R: FR, т.е.е., что ответы на низкий R: FR в первую очередь опосредуются phyB. Анализ мутантов также показал, что phyD и phyE действуют избыточно с phyB, контролируя SAS-ответы взрослых, такие как удлинение черешка и удлинение междоузлий между листьями розетки (обзор Martinez-Garcia et al., 2010; Casal, 2012).
РИСУНОК 1. Сводка фенотипов мутантных растений дикого типа, phyA и phyB , выращенных при различных условиях освещения. Фенотипы проростков (удлинение гипокотиля) и листьев (удлинение черешка, раскрытие пластинок и скручивание листьев) показаны для каждого из указанных генотипов и световых обработок. Карикатуры обобщают результаты, представленные разными авторами, упомянутыми в этом обзоре. нет данных, данные отсутствуют; WT, дикий тип.
Генетический анализ также показал, что проростки мутанта phyA демонстрируют усиленные (а не сниженные) ответы на низкий R: FR, что указывает на антагонистическое действие phyA и phyB в тени.Это особенно заметно при выращивании растений в глубокой тени. В то время как близость растительности (что приводит к умеренному R: FR) способствует аналогичному удлинению гипокотилей у проростков дикого типа и phyA , в условиях глубокой и прямой тени растительного покрова (что приводит к низким и очень низким R: FR) гипокотилям дикорастущих растений. проростки такого типа имеют тенденцию к удлинению значительно меньше, чем у проростков мутанта phyA , которые демонстрируют чрезмерное удлинение (рисунок 1; Johnson et al., 1994; Yanovsky et al., 1995; Мартинес-Гарсия и др., 2014; Поссарт и др., 2014). Следовательно, phyB деактивируется оттенком промежуточного, низкого и очень низкого R: FR, что приводит к увеличению удлинения гипокотиля. Напротив, phyA сильно активируется в основном оттенком с низким и очень низким R: FR, условиями, которые предотвращают чрезмерное удлинение проростка, вызванное дезактивацией phyB, тем самым обеспечивая оптимальное удлинение гипокотиля в глубокой тени. Следовательно, действия phyA и phyB помогают различать близость растений и прямую тень от растительного покрова (Martinez-Garcia et al., 2014).
Имитация близости и тени растений в лаборатории с помощью световой обработки: обработка с низким R: FR и обработкой в конце дня (EOD-FR)
Лабораторные условия, которые имитируют близость растений до того, как произойдет фактическое затенение полога, получили название имитируемый оттенок , а те, которые имитируют естественный прямой оттенок растений, когда происходит закрытие полога, оттенок полога (Roig-Villanova et al., 2007; Поссарт и др., 2014). Некоторые авторы, однако, используют термин имитирующий оттенок как синоним промежуточного, низкого или очень низкого R: FR обработки, т.е.е., независимо от количества используемого FR. Чтобы избежать двусмысленности и сравнить различные условия, в этой статье мы будем использовать термины (1) смоделированная тень, для обозначения любых условий, которые снижают R: FR и не были определены авторами как имитирующие близость растительности или оттенок; (2) — тень, — для тех условий, которые имитируют близость растений или растительности; и (3) оттенок навеса , к тем условиям, которые имитируют естественную прямую тень растений.Эти методы лечения выполняются в лаборатории путем применения увеличивающейся интенсивности FR к фиксированной интенсивности белого света (W), обычно обеспечиваемого люминесцентными лампами: для данного количества PAR промежуточное R: FR (около 0,50–0,30) приводит к близости Затенение, а также низкий и очень низкий R: FR (<0,29) приводит к затенению навеса (Martinez-Garcia et al., 2014). Однако эти значения R: FR могут различаться в разных лабораториях в зависимости от окна (в нм) измеренных R и FR. Как уже обсуждалось ранее, лучший способ различить или сравнить эти условия затенения - это генетический анализ дифференциального удлинения гипокотиля мутантных проростков phyA и phyB (рис. 1).
Можно подумать, что очень низкий R: FR для обработки оттенков навеса редко существует в природе: поскольку количество FR, используемого в лаборатории для дополнения W, очень велико, кажется трудным понять, как природа может производить такие высокие пропорции FR просто за счет фильтрации и дифференцированной передачи компонентов R и FR солнечного света. В этом отношении важно иметь в виду, что в богатых хлорофиллом органах, таких как листья, поглощение хлорофиллом света от ультрафиолета до R-части видимой области приводит к испусканию FR за счет флуоресценции (Thornber, 1975), свойство, которое может способствовать созданию низких и очень низких сигналов R: FR в естественных плотных куполах.Действительно, есть отчеты для куполов, в которых R: FR меньше 0,10 (см. Обзор в Smith, 1982).
Помимо добавления в W FR, альтернативный способ вызвать SAS-ответы — это обработка растений импульсом FR в течение нескольких минут в конце светового периода и непосредственно перед началом темной фазы фотопериода. Это называется обработкой в конце дня (EOD-FR) (Smith, 1982). Подобно низкому R: FR, обработка EOD-FR снижает активную форму Pfr фитохромов в последующий темный период.Эти методы обработки менее репрезентативны для сигналов естественных соседей, хотя считается, что они вызывают те оттеночные реакции, которые зависят от фотостабильных фитохромов (Johnson et al., 1994), то есть тех, которым способствует близость растительности.
Реакции избегания тени у сеянцев
Реакция рассады на близость растений и тень: в поисках простоты внутри сложности
Было показано, что растения на стадии проростков реагируют на тень на уровне гипокотилей / эпикотилей, семядолей и первичных листьев.В то время как реакция семядолей (черешка и области) на имитацию оттенка изучена плохо, удлинение гипокотиля изучалось наиболее часто и глубоко. Анализ реакции удлинения гипокотилей на моделируемую тень у Arabidopsis привел к выводу, что индукция SAS регулируется, по крайней мере частично, взаимодействием активных фитохромов с PIF (Leivar and Quail, 2011; Jeong and Choi, 2013), что приводит к быстрым изменениям в экспрессии десятков генов P HYTOCHROME R A PIDLY R EGULATED ( PAR ), которые предположительно играют важную роль в реализации SAS-ответов.Поскольку многие из этих генов PAR кодируют регуляторы транскрипции, считается, что ответы SAS являются следствием регуляции фитохромом транскрипционной сети, состоящей из нескольких типов регуляторов транскрипции, организованных в частично дублирующие функциональные модули. Генетический анализ, проведенный для измерения удлинения гипокотиля, продемонстрировал роль в регуляции SAS нескольких генов PAR . Это относится к членам HD-Zip II (например, ATHB2, ATHB4, HAT1, HAT2 и HAT3), bHLH (например, ATHB2, ATHB4, HAT1, HAT2 и HAT3), bHLH (e.g., BEE1, BIM1, HFR1, PAR1 и PIL1) и B — B O X СОДЕРЖАЩИХ (BBX) семейств белков. Анализ мутантов с измененными уровнями этих факторов позволил предположить отрицательные (HFR1, PAR1, PIL1, BBX21 и BBX22), положительные (BEE, BIM, BBX24 и BBX25) и сложные (HD-Zip II) роли в регуляции SAS. (Salter et al., 2003; Sessa et al., 2005; Roig-Villanova et al., 2006, 2007; Hornitschek et al., 2009; Sorin et al., 2009; Crocco et al., 2010; Cifuentes-Esquivel. и другие., 2013; Gangappa et al., 2013). Дальнейшие анализы также вовлекали не-PAR факторы в регуляцию ответа гипокотиля SAS, такие как HD-Zip class III и белки DELLA, репрессирующие рост (Djakovic-Petrovic et al., 2007; Brandt et al., 2012). PIF1, PIF3, PIF4, PIF5 (называемый PIFQ) и PIF7, белки семейства bHLH, которые, как известно, способствуют росту, также были идентифицированы как положительные игроки в ответе гипокотиля на SAS (Leivar et al., 2012; Li et al., 2012). В отличие от генов PAR , на экспрессию этих генов PIF не влияет смоделированный оттенок, но фосфорилированию белков PIF способствует активная форма фитохромов, что снижает стабильность PIFQ и активность связывания ДНК PIF7.Следовательно, снижение уровней активных фитохромов при низком R: FR увеличивает стабильность PIFQ и ДНК-связывающую активность PIF7 с его генами-мишенями, что приводит к усилению экспрессии PAR (Lorrain et al. , 2008; Li et al., 2012). Таким образом, кажется, что смоделированное восприятие тени быстро меняет баланс положительных и отрицательных факторов, что в конечном итоге приводит к удлинению гипокотилей.
Было показано, что обработка имитацией затемнения быстро и временно увеличивает эндогенные уровни ауксинов (Tao et al., 2008; Hornitschek et al., 2012; Ли и др., 2012; Bou-Torrent et al., 2014). SAV3 кодирует TAA1, фермент, необходимый для индуцированного оттенком биосинтеза IAA, биоактивного ауксина (Tao et al., 2008). Гены YUC кодируют белки, подобные флавинмонооксигеназе, которые катализируют лимитирующую стадию биосинтеза ИУК. Действительно, семейства TAA и YUC функционируют в одном и том же пути биосинтеза ауксина, известном как путь индол-2-пировиноградной кислоты (IPA) (рис.2), который продуцирует ИУК, необходимую для развития растений (Mashiguchi et al., 2011; Чжао, 2014). Было показано, что PIF4, PIF5 и PIF7 непосредственно регулируют индуцированную оттенком экспрессию некоторых генов YUC ; кроме того, опосредованное оттенком увеличение ауксина было значительно снижено как у мутантных проростков pif4pif5 , так и у pif7 . Эти результаты привели к предположению, что PIF4, PIF5 и PIF7 могут напрямую контролировать биосинтез ауксина. PIF также напрямую регулируют гены, чувствительные к ауксину, такие как INDOLE-3-ACETIC ACID INDUCIBLE 29, ( IAA29, ), что также указывает на возможное участие PIF в модуляции чувствительности к ауксину (Hornitschek et al., 2012; Ли и др., 2012). Генетический анализ показывает, что факторы транскрипции BIM также могут играть прямую или косвенную роль в синтезе ауксина посредством контроля экспрессии YUC (Cifuentes-Esquivel et al., 2013). Кроме того, несколько факторов PAR, такие как белки ATBh5, PAR1, BIM и BEE, обеспечивают точки входа, с помощью которых интегрируются сети, регулируемые оттенком и ауксином (Sorin et al., 2009; Bou-Torrent et al., 2014) . Таким образом, существует множество доказательств участия нескольких факторов в контроле уровней ауксина и чувствительности в ответе на удлинение гипокотиля.Однако, как ауксин и другие гормоны участвуют в контроле удлинения гипокотилей, индуцированного оттенком, в настоящее время неясно.
РИСУНОК 2. Схема, поясняющая, как действие фитохромов может регулировать ауксины (биосинтез, транспорт и чувствительность) в ответ проростков и листьев взрослых растений на близость растений. Восприятие оттенка интегрируется с ауксинами на нескольких уровнях, влияя как на биосинтез ауксина, так и на его чувствительность. Неизвестно, влияет ли восприятие тени также на транспорт ауксина.Две стадии биосинтеза ауксина (заключенные в черные пунктирные квадраты и представленные толстыми пустыми стрелками) указаны вместе с участвующими ферментами (SAV3 / TAA1 и YUC). Транспорт ауксина обозначен серым цветом, а чувствительность к ауксину — фиолетовым. Черные и фиолетовые линии указывают на то, что роль известных регуляторов может быть прямой (непрерывной) или косвенной (прерывистой), а также положительной (стрелки) или отрицательной (плоские концы). Серые стрелки указывают направление транспорта ИУК в проростках или листьях, тогда как синяя стрелка указывает сигнал от верхушки побега (область, ограниченная красным квадратом) к семядолям, предложенным Nito et al.(2015). Trp, тритофан; IPA, индол-2-пировиноградная кислота; ИУК, индол-3-уксусная кислота.
Физическое разделение между участками восприятия тени и ответа: межорганная коммуникация
Существует дополнительный пространственный уровень регуляции ответов проростков на SAS. Физическое разделение между участками восприятия тени и ответной реакции подчеркивается множеством экспериментальных подходов к различным видам, которые использовали локализованное теневое облучение (EOD-FR или низкий R: FR) семядолей (или первичных листьев, как мы увидим позже). в сочетании с молекулярным или физиологическим анализом отвечающих стволов (гипокотилей или эпикотилей; обзор Bou-Torrent et al., 2008).
Таким образом, подробный анализ проростков Brassica rapa показал, что восприятие тени семядолями, местом световосприятия этого сигнала у проростков, запускает локальный синтез ИУК, которая затем транспортируется в гипокотиль. Там ауксин индуцирует повышающую регуляцию связанных с ауксином генов, связанных с ответом гипокотиля на SAS. Вместе эти результаты показывают, что генерируемый семядолями ауксин регулирует удлинение гипокотилей (Procko et al., 2014). Органоспецифический транскриптомный анализ с использованием микропрепаратов, приготовленных из верхней части гипокотилей (называемых образцами верхушки побега) и семядолей молодых проростков Arabidopsis , дифференцированно облученных с EOD-FR или без него, показал, что реакция на оттенок с точки зрения изменения в экспрессии генов в верхушке побега более заметны, чем в семядолях, вероятно, отражая более высокое разнообразие типов клеток в образцах верхушки побега.Обнаружение специфических и общих генов, индуцированных EOD-FR, в обоих типах образцов предполагает существование тканеспецифичных и общих регуляторных компонентов, участвующих в реализации SAS-ответов в различных тканях. Кроме того, локализованное облучение прожектором EOD-FR также показало, что экспрессия генов, индуцированная EOD-FR, зависит как от автономных органов (т.е. HFR1 и ATHB2 ), так и от межорганных механизмов, которые включают роль ауксинов и PIF. В самом деле, семядоли являются основным участком восприятия оттенка, который контролирует несколько генов, чувствительных к верхушке, включая те, которые зависят от ауксина и / или PIF7.Это согласуется с известной ролью PIF7, преимущественно регулирующего SAS-ответ проростков, контролируя биосинтез ауксина в семядолях (Рисунок 2). Однако сигнал оттенка, воспринимаемый на верхушке побега, также контролирует экспрессию генов в семядолях (т.е. YUC2 ), что свидетельствует о неожиданной двунаправленной коммуникации между этими частями проростков (Nito et al., 2015). Хотя физиологическая значимость двусторонней коммуникации неясна, кажется вероятным, что это часть механизма обратной связи для координации реакции различных органов проростков.
Реакция избегания тени у взрослых растений
У проростков низкий R: FR способствует удлинению черешка семядолей у Arabidopsis и ингибирует рост семядолей у B. rapa (Roig-Villanova et al., 2007; Procko et al., 2014). У растений, которые имеют привычку к росту без розетки, таких как вигна, табак или горчица, удлинение междоузлий также сильно стимулируется в ответ на различные обработки, имитирующие оттенок (Casal and Smith, 1988; Martinez-Garcia et al., 2000; Pierik et al., 2004). Еще более сложным образом, чем у проростков, Arabidopsis демонстрирует широкий диапазон реакций на тень во взрослых тканях, таких как стимулирование удлинения черешка в листьях розетки (обычно вместе с ингибированием расширения листовой пластинки), стимулирование цветения. длина стебля соцветия вместе с подавлением разрастания пазушных почек (Finlayson et al., 2010; Gonzalez-Grandio et al., 2013). В этом разделе будут рассмотрены только реакции листа, включая влияние на полярность листа.Мы также обсудим роль тени на репродуктивном развитии.
Удлинение черешка, разрастание листьев и скручивание листьев
Лист Arabidopsis состоит из пластинки (или листовой пластинки) и черешка (рис. 3А). Черешок помещает пластинку листа в положение, которое лучше всего подходит для приема света. Следовательно, регуляция удлинения черешка кажется важной для максимального фотосинтеза. Одним из наиболее изученных SAS-ответов розеточных листьев Arabidopsis является удлинение черешка, которому способствуют световые обработки, имитирующие близость растительности или оттенок полога, такие как низкий R: FR и EOD-FR (Kozuka et al., 2005; Сасидхаран и др., 2010; Рисунки 1 и 3А). Генетический анализ показывает, что удлинение черешков листьев регулируется фотостабильными фитохромами: мутант phyB имеет более длинные черешки, чем у дикого типа (рис. 1), а мутанты phyAphyBphyD и phyAphyBphyE имеют более длинные черешки, чем мутант phyAphy. (Рид и др., 1993; Девлин и др., 1998). Следовательно, стимуляция роста черешка с помощью сигналов тени опосредуется главным образом phyB с участием phyD и phyE (Casal, 2012).
РИСУНОК 3. Взрослые листья проявляют двойную реакцию на близость растений и тень. (A) Рисунок , представляющий две части, которые образуют взрослый лист Arabidopsis , и их дифференциальную реакцию роста на EOD-FR или имитацию затенения: стимулирование роста длины черешка и ингибирование роста лопастей. (B) Диаграмма Венна, иллюстрирующая различия в экспрессии генов, индуцированных EOD-FR в лопатках и черешках, позволяет идентифицировать основной набор генов, которые индуцируются в обеих тканях.Данные взяты из доступных данных микрочипов (Sasidharan and Pierik, 2010).
Для роста и удлинения органов растения требуется, чтобы обычно жесткая клеточная стенка ослаблялась и, таким образом, позволяла продолжать расширение. XTHs представляют собой хорошо охарактеризованные белки, модифицирующие клеточную стенку, которые участвуют в расширении клеток (Rose et al., 2002). Действительно, удлинение черешка связано с увеличением активности XTH, который ферментативно действует (гидролиз и / или трансглюкозилирование) на сеть целлюлозы и гемицеллюлозы (ксилоглюкан у большинства эвдикотов) и приводит к необходимому разрыхлению клеточной стенки.В Arabidopsis XTH кодируются большим мультигенным семейством, и экспрессия некоторых из них ( XTH9 , XTh25 / XTR7 , XTh26 , XTh27 и XTh29 ) регулируется с повышением. по оттенку. Однако только нокаут-мутанты для XTh25 и XTh27 имели пониженные или отсутствующие SAS-ответы черешка. Вместе эти результаты подтверждают функциональную роль некоторых XTH в реструктуризации клеточной стенки, необходимой для клеточной экспансии, которая способствует быстрому удлинению черешка во время SAS (Sasidharan et al., 2010).
В дополнение к стимулированию удлинения черешка, рост и расширение листовых пластинок подавляется как в условиях имитации тени, так и при обработке EOD-FR (Рисунки 1 и 3A; Kozuka et al., 2010; de Wit et al., 2015) . Два антагонистических эффекта тени на рост листьев, скорее всего, сопутствуют и взаимосвязаны (рис. 3А). Это говорит о том, что, когда листья находятся в непосредственной близости или в тени навеса, рост вкладывается в удлинение черешка за счет расширения лезвия.Обработка EOD-FR сильнее способствует удлинению базальной части черешка, хотя и базальная, и апикальная части этого органа удлиняются. Точно так же EOD-FR подавлял в основном расширение средней части листовых пластинок (рис. 3А). На стадии анализа листа оба эффекта происходили за счет дифференциального контроля удлинения клеток с незначительным вкладом клеточного деления (Kozuka et al., 2005). Другие авторы показали, что уменьшенная площадь лопастей первого листа, полностью выросшего в тени навеса, и пониженная интенсивность света вызваны уменьшением пролиферации клеток, а не размером клеток (Carabelli et al., 2007). Эти противоположные наблюдения могут объяснить различную обработку оттенка и возраст листьев, на которые была нанесена тень в разных лабораториях, и предполагают, что воздействие близости или тени навеса может влиять в первую очередь на деление клеток на ранних стадиях развития листа и, в основном, на рост клеток уже в сформированные листья. Растения с мутантом phyB демонстрируют удлиненные черешки и более мелкие листовые пластинки по сравнению с растениями дикого типа, фенотип, который указывает на роль phyB в регуляции этих признаков противоположным образом (рисунок 1; Robson et al., 1993; Kozuka et al., 2005).
Таким образом, кажется, что оттенок одинаково влияет на характер роста фотосинтетических органов (то есть подавляет рост листовой пластинки и семядолей) и поддерживающих органов (то есть способствует удлинению черешка листа и гипокотиля). Неизвестно, контролируют ли регуляторы удлинения гипокотиля, индуцированного оттенком, также удлинение черешка, индуцированное оттенком. Доступные транскриптомные данные предполагают существование некоторых общих регуляторов (Kozuka et al., 2010). Анализ микрочипов показал, что многие гены, реагирующие на ауксин и брассиностероиды (BR), проявляют измененную экспрессию в ответ на EOD-FR как в листовой пластинке, так и в черешке. Действительно, 52 и 81 ген, отвечающий за ауксин, включая IAA , Gh4 и SMALL AUXIN UPREGULATED RNA ( SAUR ) семейств, были активированы EOD-FR в пластинке листа (из общего числа 109 генов) и черешка (из 168 генов) соответственно. Гены, которые дважды контролируются ауксином и BR, были чрезмерно представлены в генах, индуцированных EOD-FR, также в листовых пластинках и черешках (Kozuka et al., 2010). Точно так же быстрые изменения в экспрессии нескольких генов, чувствительных к ауксину и BR, наблюдались у проростков, подвергшихся воздействию искусственной тени (Bou-Torrent et al., 2014). Среди генов, индуцированных обработкой EOD-FR на черешках и / или лопастях, некоторые из них кодируют регуляторы транскрипции, которые, как известно, играют важную роль в реакции проростков SAS, такие как ATHB2 , HAT2 , HAT3 , HFR1. , PAR1 , BEE1 и BIM1 (рисунок 3; дополнительная таблица S1).Некоторые из этих генов напрямую регулируются PIF (например, ATHB2 , HFR1 и PAR1 ), что позволяет предположить, что эти факторы также играют регулирующую роль в росте и разрастании листьев, индуцированных затенением. Хотя еще нет экспериментальных данных, подтверждающих эту возможность, одиночные или множественные мутанты с дефицитом PIF , такие как pif4 и двойной pif4pif5 , демонстрируют маленькие и компактные розетки; в соответствии с этим, растения со сверхэкспрессией PIF4 и PIF5 демонстрируют расширенные розетки с сильно удлиненными черешковыми листьями при различных условиях освещения или температур (Pierik et al., 2004; Lorrain et al., 2008; Sasidharan et al., 2010). Недавние достижения показали, что PIF7 играет преобладающую роль в регуляции удлинения черешка и расширения листовой пластинки в ответ на низкий R: FR, тогда как PIF4, PIF5 и HFR1 играют незначительную роль или не играют никакой роли в этих ответах листа. Кроме того, уровни ауксина индуцируются в листовой пластинке вскоре после воздействия низкого R: FR, всплеска, который отсутствует у мутанта pif7 (de Wit et al., 2015). Эти данные подтверждают существование как общих (PIF7), так и уникальных (PIF4, PIF5, HFR1) регуляторных компонентов и / или механизмов SAS на разных стадиях развития, т.е.г., гипокотиль и рост листьев.
Восприятие тени розеточными растениями, такими как Arabidopsis , листья которых находятся близко к земле, обычно приводит к изменению положения листьев, что помогает избежать затенения, создаваемого соседними растениями. Изгиб листьев вверх, вызванный более быстрым ростом их нижней, чем верхней стороны, называется гипонастией. Гипонастический ответ на низкий R: FR сильно снижен у ряда мутантов ауксина, таких как sav3 , который кодирует фермент, участвующий в быстром повышении эндогенных уровней ИУК, необходимых для индуцированного оттенком удлинения гипокотиля (рис. 2). и pin3 , который кодирует белок, который контролирует направление и скорость клеточного потока ауксина, указывая тем самым, что гипонастический ответ листа на моделируемую тень требует интактной передачи сигналов ауксина (Casal, 2012).
Близость или оттенок навеса, по-видимому, также влияет на дополнительный аспект развития листовой пластинки: скручивание или уплощение. Считается, что регулировка плоскостности листьев способствует эффективному поглощению света при слабом освещении, в условиях, сопровождающих тень навеса. При анализе растений phyB дикого типа и растений , выращенных в условиях непрерывного W, было обнаружено, что у растений дикого типа четвертые листья были слегка загнуты вниз, тогда как у растений phyB листья были плоскими.В трансгенных линиях, сверхэкспрессирующих PHYB , листья были более сильно скручены, чем у дикого типа, что указывает на то, что phyB ингибирует сплющивание листьев (Kozuka et al., 2013). Кроме того, у дикого типа уплощению листьев способствовала обработка EOD-FR, которая быстро устраняла активный фитохром Pfr. В то время как phyB демонстрировал очень плоские листья без EOD-FR и не реагировал на EOD-FR, фенотип уже свернутых листьев у растений со сверхэкспрессией PHYB был существенно подавлен EOD-FR.Таким образом, был сделан вывод, что активная форма Pfr phyB способствует скручиванию листьев, процессу, который может включать регуляцию реакции на ауксин в этом органе (Kozuka et al., 2013).
Скручивание листа, скорее всего, вызвано неравномерным распространением эпидермальных клеток на адаксиальной (верхней) и абаксиальной (нижней) сторонах листа, хотя информации о влиянии тени на площадь эпидермальных клеток на этих сторонах листьев нет. Интересно обнаружить, что мутанты с измененными уровнями ATHB4 и / или HAT3 , два гена паралогов, кодирующие факторы транскрипции класса II HD-Zip и идентифицированные как регуляторы ответа удлинения гипокотилей SAS (Sorin et al., 2009), действительно показывают роль в контроле дорсо-вентрального паттерна листьев. В то время как двойные мутантные растения athb4 hat3 приводят к сильно абаксиализированным листьям, сверхэкспрессия HAT3 приводит к адаксиализированному развитию листьев, что визуализируется сильным загибанием вверх листовых пластинок, фенотипом, вызванным чрезмерным размножением абаксиальных тканей в листьях. (Bou-Torrent et al., 2012). Поскольку длительное пребывание в тени приводит к появлению листьев со сплющенными лопастями, эти результаты позволяют предположить, что ATHB4 и HAT3 , как было показано, являются функционально избыточными (Sorin et al., 2009), может быть частью механизмов, регулирующих этот SAS-ответ у взрослых листьев.
Таким образом, на стадии розетки световые сигналы затенения способствуют перемещению листьев вверх и удлинению черешка; обычно они также уменьшают расширение листовой пластинки и усиливают уплощение листьев (рис. 1). Комбинация этих реакций помещает листовые пластинки вдали от тени соседей, что, вероятно, усиливает фотосинтетическую активность растения. Текущие данные подтверждают, что некоторые регуляторы этих ответов могут быть общими с таковыми у проростков, несмотря на структурные различия между задействованными органами.Совсем недавно было также высказано предположение, что растения Arabidopsis переориентируют свои листья после распознавания родственников-соседей (Crepy and Casal, 2015), хотя есть некоторые разногласия относительно того, является ли это скорее случаем соответствия фенотипу (Crepy and Casal, 2016; Тиль-Боттро и де Вильмерёй, 2016). Благотворное влияние этих перестроек на приспособленность растений путем ослабления жесткой конкуренции за свет открывает интересный предмет для изучения.
ОблучениеSpotlight EOD-FR, примененное отдельно к листовой пластинке и черешку, показало, что фоточувствительным участком для регуляции удлинения черешка была пластинка листа, а не сам черешок.Молекула-кандидат в мобильный сигнал — это опять же ауксин. Анализ экспрессии Gh4.3 ( At2g23170 ) и IAA6 ( At1g52830 ), двух ауксин-чувствительных генов в черешке, идентифицированных ранее как индуцируемых после обработки EOD-FR всего растения, показал, что их экспрессия находилась под контролем фитохрома в листовой пластинке; То есть свет, направленный на листовую пластинку, эффективно индуцировал экспрессию генов не только в листовой пластинке, но и на черешке.Напротив, облучение черешка вообще не было эффективным для индукции экспрессии этих двух генов в самом черешке. Вместе эти простые и элегантные эксперименты показали, что действие фитохромов в листовой пластинке контролирует экспрессию генов и ответы удлинения черешка (Kozuka et al., 2010). Транспорт ауксина от лезвия к черешку, по-видимому, играет роль в этой реакции удлинения, поскольку ингибитор транспорта ауксина NPA специфически подавляет удлинение черешка в ответ на EOD-FR или на низкий R: FR (Pierik et al., 2009; Kozuka et al., 2010), даже когда NPA применяли локально только в месте соединения черешка и лопасти (de Wit et al., 2015). Совсем недавно было показано, что обработка низким R: FR быстро увеличивает уровни ИУК в листовой пластинке, процесс зависит от PIF7 (de Wit et al., 2015), что позволяет предположить, что имеет место индуцированная низким R: FR продукция ауксина. в основном в листовой пластине и затем будет транспортироваться к черешку, сообщению между пластинкой и черешком, которое может быть аналогично сообщению между семядолями и гипокотилем (Hornitschek et al., 2012; Ли и др., 2012; Bou-Torrent et al., 2014). Однако аналогичная кинетика уровней ауксина в лопастях и черешках растений с низким R: FR не полностью соответствует этой модели, что позволяет предположить, что другие аспекты, такие как изменения чувствительности к ауксину, также могут иметь роль в этом SAS-ответе. .
SAS Responses во время размножения растений: выбор Софи?
Растения Arabidopsis , выращенные в фотопериодических условиях, в которых светлая фаза имеет низкий R: FR, демонстрируют заметное сокращение времени цветения.Поскольку этот ответ был недавно подробно описан (Casal, 2012), он не будет здесь обсуждаться. Ускоренное цветение растений Arabidopsis было связано с уменьшенным завязыванием семян, усеченным развитием плодов и часто с серьезным снижением всхожести полученных семян (Smith and Whitelam, 1997), хотя была очень значительная генетическая изменчивость реакции прорастания. найдено (Dechaine et al., 2009). Однако очень мало информации об этих ответах SAS у Arabidopsis или родственных видов.Недавно было описано, что взрослых растений B. rapa , выращенных в искусственной тени, имели заметно более длинные стручки, давали меньше зрелых семян на стручок, и эти семена были меньше (Procko et al., 2014). Arabidopsis растений, выращенных в тех же условиях, значительно снизили урожай семян с одного растения (Procko et al., 2014). В условиях W мутантные растения с дефицитом phyB B. rapa (обозначенные как ein194 ) демонстрировали пониженное количество семян на стручок по сравнению с растениями дикого типа, что позволяет предположить, что некоторые из этих фенотипов частично определяются передачей сигналов phyB.Активность PhyB, по-видимому, не играет значительной роли в определении других исследованных фенотипов, таких как длина стручка и масса семян (Procko et al., 2014). В Arabidopsis начинают выясняться эндогенные факторы, определяющие количество семяпочек / семян [обзор (Cucinotta et al., 2014)]. Однако из-за плохой характеристики оттенков у взрослых репродуктивных стадий Arabidopsis , практически отсутствует информация о регуляции этого сигнала окружающей среды.
Уровни транскриптов нескольких генов PAR , таких как PHYB , HFR1 , PIL1 , ATHB4 и HAT3 , были значительно увеличены в условиях низкого R: FR в соцветиях, что указывает на то, что репродуктивные ткани воспринимают и реагируют (по крайней мере, на молекулярном уровне) на близость растений или тень (Reymond et al., 2012). В проростках экспрессия PIL1 и HFR1 напрямую регулируется действием некоторых PIF в ответ на имитацию тени (Hornitschek et al., 2012). Следовательно, будет интересно выяснить, играют ли PIFs также роль в регуляции упомянутых SAS-ответов в репродуктивных тканях.
В целом считается, что ответы SAS перенаправляют фотоассимиляты в сторону удлинения за счет запасающих органов. На репродуктивных стадиях, в которых не происходит значительного удлинения, воздействие близости растений или тени навеса ставит растения перед другой дилеммой: вкладывать ли деньги в производство большого количества семян, многие из которых могут оказаться нежизнеспособными из-за недостаточности фотоассимилятов. или в ограниченном количестве жизнеспособных семян.В этом выборе Софи растения, похоже, отдают предпочтение второй стратегии. Поиск задействованных регулирующих компонентов и механизмов может помочь в разработке новых способов улучшения сельскохозяйственных культур в грядущие и трудные времена.
Заключительные замечания
В этом обзоре мы подчеркнули идею о том, что для того, чтобы приспособить рост растений к условиям тени, ответы могут варьироваться в зависимости от органа, стадии развития и / или рассматриваемых видов. Эти исследования были сосредоточены в основном на Arabidopsis и родственных двудольных.У однодольных растений большинство проанализированных реакций на тень относятся к изменениям в архитектуре растения, которые сильно снижают урожайность. Действительно, как и в случае Arabidopsis и других двудольных растений, устойчивые реакции SAS у трав могут отвлекать ресурсы на удлинение стебля за счет производства зерна (обзор Warnasooriya and Brutnell, 2014).
Несмотря на усилия, предпринятые многими лабораториями в течение последних десятилетий для понимания SAS (обзор в Martinez-Garcia et al., 2010; Casal, 2012; Pierik and de Wit, 2014), все еще мало информации об этих ответах у взрослых. растения.Существует, например, особый недостаток знаний о световосприимчивом участке тени в SAS-ответах в репродуктивных тканях. Что касается усиления оттенка цветения у Arabidopsis , представляется вероятным, что место восприятия тени (листья) отделено от места действия (апикальная меристема побега), как это, как известно, происходит в зависимости от фотопериода. индукция цветения. Однако нет информации об этом аспекте в SAS-ответах репродуктивных тканей, хотя, как уже упоминалось, ткани соцветий воспринимают близость растений и реагируют на нее на молекулярном уровне (Reymond et al., 2012). Учитывая, что у Arabidopsis , когда соцветия полностью разрастаются, розеточные листья уже стареют, кажется вероятным, что для этих ответов место восприятия и действия совпадают. Таким образом, будущие исследования по характеристике SAS-ответов на стадиях взрослого растения, а также его пространственной и генетической регуляции внесут большой вклад в понимание этого важного адаптивного признака, имеющего решающее значение для урожайности сельскохозяйственных культур.
Авторские взносы
Рукопись написалиIR-V и JFM-G.
Финансирование
IR-V имеет контракт с Беатриу де Пинос от Agència de Gestió D’Ajuts Universitaris i de Recerca (AGAUR, Generalitat de Catalunya). Наше исследование поддерживается грантами от испанского Министерства экономики и конкуренции (BIO2014-59895-P) и AGAUR (2014-SGR447 и Xarba) для JFM-G.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарность
Мы очень благодарны Бриардо Льоренте за критические комментарии к рукописи, а также членам лаборатории (Мария Хосе Молина и Санди Паулишич) за их комментарии и предложения.
Дополнительные материалы
Дополнительные материалы к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/article/10.3389/fpls.2016.00236
Сокращения
ATHB, ARABIDOPSIS THALIANA HOMEOBOX; BBX, B-BOX, СОДЕРЖАЩИЕ белки; ПЧЕЛА, BR РАСШИРЕННОЕ ВЫРАЖЕНИЕ; bHLH, основная спираль-петля-спираль; BIM, BES1-ВЗАИМОДЕЙСТВУЮЩИЙ MYC-LIKE; EOD-FR, конец дня — дальний красный; FR — дальний красный свет; ШАПКА, HOMEOBOX FROM ARABIDOPSIS THALIANA ; HD-Zip II, гомедоменная лейциновая молния класса II; HFR1, ДЛИННЫЙ ГИПОКОТИЛ В ДАЛЬНЕМ-КРАСНОМ1; ИУК, индол-3-уксусная кислота; ген IAA , ген IAA INDUCIBLE ; IPA, индол-2-пировиноградная кислота; NPA, 1-N-нафтилфталаминовая кислота; PAR — фотосинтетическая активная радиация; PAR гена, БЫСТРО РЕГУЛИРУЕМЫЙ ФИТОХРОМ гена; PHY, ФИТОХРОМ; СДС, ФАКТОР ВЗАИМОДЕЙСТВИЯ С ФИТОХРОМОМ; PIFQ, квартет PIF; PIL1, PIF3-LIKE 1; КОНТАКТ 3, ФОРМОВАННЫЙ КОНТАКТ 3; Pfr, активная форма фитохромов; Pr, неактивная форма фитохромов; R — красный свет; R: FR, отношение R к FR; САС, синдром избегания тени; SAUR гена, МАЛЕНЬКАЯ АУКСИН-УПРЕГУЛИРОВАННАЯ РНК гена; SAV3, ИЗБЕЖАНИЕ ТЕНЕЙ 3; TAA1, ТРИПТОФАНАМИНОТРАНСФЕРАЗА ARABIDOPSIS 1; W, белый свет; XTHs, ксилоглюкан-эндотрансглюкозилаза / гидролазы; гена YUC , гена YUCCA .
Список литературы
Бу-Торрент, Дж., Галстян, А., Галлеми, М., Сифуэнтес-Эскивель, Н., Молина-Контрерас, М. Дж., Салла-Мартрет, М., и др. (2014). Восприятие близости растений динамически модулирует уровень гормонов и чувствительность у Arabidopsis . J. Exp. Бот. 65, 2937–2947. DOI: 10.1093 / jxb / eru083
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бу-Торрент, Дж., Салла-Мартрет, М., Брандт, Р., Мусиелак, Т., Палауки, Дж.К., Мартинес-Гарсия, Дж. Ф. и др. (2012). ATHB4 и HAT3, два транскрипционных фактора HD-ZIP класса II, контролируют развитие листьев у Arabidopsis . Завод Сигнал. Behav. 7, 1382–1387. DOI: 10.4161 / psb.21824
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Брандт Р., Салла-Мартрет М., Бу-Торрент Дж., Мусиелак Т., Шталь М., Ланц К. и др. (2012). Полногеномный анализ сайтов связывания REVOLUTA выявляет связь между формированием паттерна листьев и ответами роста, опосредованными светом. Plant J. 72, 31–42. DOI: 10.1111 / j.1365-313X.2012.05049.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Карабелли М., Поссенти М., Сесса Г., Чиолфи А., Сасси М., Морелли Г. и др. (2007). Тень навеса вызывает быструю и временную остановку развития листьев за счет индуцированной ауксином активности цитокининоксидазы. Genes Dev. 21, 1863–1868. DOI: 10.1101 / gad.432607
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Casal, J.Дж. И Смит Х. (1988). Устойчивые эффекты изменений статуса фитохромов на рост междоузлий у выращенной на свету горчицы: возникновение, кинетика и локус восприятия. Planta 175, 214–220. DOI: 10.1007 / BF00392430
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Cifuentes-Esquivel, N., Bou-Torrent, J., Galstyan, A., Gallemi, M., Sessa, G., Salla Martret, M., et al. (2013). Белки bHLH BEE и BIM положительно модулируют синдром избегания тени у проростков Arabidopsis . Plant J. 75, 989–1002. DOI: 10.1111 / tpj.12264
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Крокко, К. Д., Холм, М., Яновский, М. Дж., И Ботто, Дж. Ф. (2010). AtBBX21 и COP1 генетически взаимодействуют в регуляции избегания тени. Plant J. 64, 551–562. DOI: 10.1111 / j.1365-313X.2010.04360.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
де Вит, М., Юнг, К., Фанкхаузер, К. (2015). Контрастные реакции роста в пластинке и черешке во время обнаружения соседей зависят от дифференциальной реакции на ауксин, а не от различных уровней ауксина. New Phytol. 208, 198–209. DOI: 10.1111 / nph.13449
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дечайн, Дж. М., Гарднер, Г., и Вайниг, К. (2009). Фитохромы по-разному регулируют реакцию прорастания семян на световые и температурные сигналы во время созревания семян. Plant Cell Environ. 32, 1297–1309. DOI: 10.1111 / j.1365-3040.2009.01998.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Девлин, П.Ф., Патель, С. Р., и Уителам, Г. К. (1998). Фитохром E влияет на удлинение междоузлий и время цветения у Arabidopsis . Растительная клетка 10, 1479–1487. DOI: 10.1105 / tpc.10.9.1479
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джакович-Петрович, Т., де Вит, М., Военек, Л.А., и Пиерик, Р. (2007). Белок DELLA участвует в ответных реакциях роста на сигналы растительного покрова. Plant J. 51, 117–126. DOI: 10.1111 / j.1365-313X.2007.03122.х
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Финлейсон, С. А., Кришнаредди, С. Р., Кебром, Т. Х., Касал, Дж. Дж. (2010). Фитохромная регуляция ветвления у Arabidopsis . Plant Physiol. 152, 1914–1927. DOI: 10.1104 / стр.109.148833
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гангаппа, С. Н., Крокко, К. Д., Йоханссон, Х., Датта, С., Хеттиараччи, К., Холм, М., и др. (2013). Белок B-BOX Arabidopsis BBX25 взаимодействует с HY5, негативно регулируя экспрессию BBX22 и подавляя фотоморфогенез проростков. Растительная клетка 25, 1243–1257. DOI: 10.1105 / tpc.113.109751
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гоммерс, К. М., Виссер, Э. Дж., Сент-Онге, К. Р., Военек, Л. А., и Пиерик, Р. (2013). Теневыносливость: при высоком росте — не вариант. Trends Plant Sci. 18, 65–71. DOI: 10.1016 / j.tplants.2012.09.008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Gonzalez-Grandio, E., Poza-Carrion, C., Sorzano, C.О., и Кубас, П. (2013). BRANCHED1 способствует покою пазушных почек в ответ на тень у Arabidopsis . Растительная клетка 25, 834–850. DOI: 10.1105 / tpc.112.108480
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хорницчек, П., Конен, М. В., Лоррен, С., Ружмонт, Дж., Юнг, К., Лопес-Видриеро, И. и др. (2012). Взаимодействующие с фитохромом факторы 4 и 5 контролируют рост проростков в изменяющихся условиях освещения, напрямую контролируя передачу сигналов ауксина. Plant J. 71, 699–711. DOI: 10.1111 / j.1365-313X.2012.05033.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хорницчек, П., Лоррен, С., Зоте, В., Михиелин, О., Фанкхаузер, К. (2009). Ингибирование реакции избегания тени за счет образования не связывающихся с ДНК гетеродимеров bHLH. EMBO J. 28, 3893–3902. DOI: 10.1038 / emboj.2009.306
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джонсон, Э., Брэдли, М., Харберд, Н.П., и Уайтелам, Г.С. (1994). Фотоответы выращенных на свету мутантов phyA Arabidopsis (фитохром А необходим для восприятия увеличения продолжительности светового дня). Plant Physiol. 105, 141–149.
PubMed Аннотация | Google Scholar
Кодзука Т., Хоригути Г., Ким Г. Т., Огиши М., Сакаи Т. и Цукая Х. (2005). Различные реакции роста листовой пластинки Arabidopsis thaliana и черешка во время избегания тени регулируются фоторецепторами и сахаром. Physiol растительных клеток. 46, 213–223. DOI: 10.1093 / pcp / pci016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кодзука Т., Кобаяси Дж., Хоригути Г., Демура Т., Сакакибара Х., Цукая Х. и др. (2010). Вовлечение ауксина и брассиностероида в регуляцию удлинения черешка в тени. Plant Physiol. 153, 1608–1618. DOI: 10.1104 / стр.110.156802
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Козука, Т., Суэцугу, Н., Вада, М., и Нагатани, А. (2013). Антагонистическая регуляция уплощения листьев фитохромом B и фототропином у Arabidopsis thaliana . Physiol растительных клеток. 54, 69–79. DOI: 10,1093 / pcp / шт134
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лейвар П., Тепперман Дж. М., Кон, М. М., Монте, Э., Аль-Сади, Б., Эриксон, Э. и др. (2012). Динамический антагонизм между фитохромами и основными факторами семейства PIF спираль-петля-спираль индуцирует избирательные реципрокные ответы на свет и тень в быстро реагирующей транскрипционной сети Arabidopsis . Растительная клетка 24, 1398–1419. DOI: 10.1105 / tpc.112.095711
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Li, L., Ljung, K., Breton, G., Schmitz, R.J., Pruneda-Paz, J., Cowing-Zitron, C., et al. (2012). Связь возбуждения фоторецепторов с изменениями в архитектуре растений. Genes Dev. 26, 785–790. DOI: 10.1101 / gad.187849.112
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лоррен, С., Аллен, Т., Дуэк, П. Д., Whitelam, G.C. и Fankhauser, C. (2008). Опосредованное фитохромом ингибирование избегания тени включает деградацию стимулирующих рост факторов транскрипции bHLH. Plant J. 53, 312–323. DOI: 10.1111 / j.1365-313X.2007.03341.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мартинес-Гарсия, Дж. Ф., Галлеми, М., Молина-Контрерас, М. Дж., Льоренте, Б., Бевилаква, М. Р., Куэйл, П. Х. (2014). Синдром избегания тени у Arabidopsis : антагонистическая роль фитохрома А и В различает близость растительности и оттенок полога. PLoS ONE 9: e109275. DOI: 10.1371 / journal.pone.0109275
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Мартинес-Гарсия, Дж. Ф., Галстян, А., Салла-Мартрет, М., Сифуэнтес-Эскивель, Н., Галлеми, М., и Бу-Торрент, Дж. (2010). Регуляторные компоненты синдрома избегания тени. Adv. Бот. Res. 53, 65–116. DOI: 10.1016 / S0065-2296 (10) 53003-9
CrossRef Полный текст | Google Scholar
Мартинес-Гарсия, Х. Ф., Сантес, К.М., и Гарсия-Мартинес, Дж. Л. (2000). Дальнее красное облучение в конце дня увеличивает содержание гиббереллина A1 в Copwa ( Vigna sinensis L.) за счет снижения его инактивации. Physiol. Растение. 108, 426–434. DOI: 10.1034 / j.1399-3054.2000.t01-1-100413.x
CrossRef Полный текст | Google Scholar
Машигучи К., Танака К., Сакаи Т., Сугавара С., Кавайде Х., Нацуме М. и др. (2011). Основной путь биосинтеза ауксина у Arabidopsis . Proc.Natl. Акад. Sci. США 108, 18512–18517. DOI: 10.1073 / pnas.1108434108
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Нито К., Кадзияма Т., Унтен-Кобаяши Дж., Фуджи А., Мочизуки Н., Камбара Х. и др. (2015). Пространственная регуляция ответа экспрессии гена на оттенок у проростков Arabidopsis . Physiol растительных клеток. 56, 1306–1319. DOI: 10.1093 / pcp / pcv057
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пиерик, Р., Куппенс, М. Л., Военек, Л. А., и Виссер, Э. Дж. (2004). Взаимодействие между этиленом и гиббереллинами в реакции избегания оттенка, опосредованной фитохромом, в табаке. Plant Physiol. 136, 2928–2936. DOI: 10.1104 / стр.104.045120
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пиерик, Р., Джакович-Петрович, Т., Кеускамп, Д. Х., де Вит, М., и Военек, Л. А. (2009). Ауксин и этилен регулируют элонгационные ответы на сигналы близости соседей независимо от гиббереллина и белков делла в Arabidopsis . Plant Physiol. 149, 1701–1712. DOI: 10.1104 / стр.108.133496
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Поссарт А., Флек К. и Хилтбруннер А. (2014). Излучение (дальний красный) свет на механизмы и реакции фитохромов у наземных растений. Plant Sci. 21, 36–46. DOI: 10.1016 / j.plantsci.2013.11.013
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Procko, C., Crenshaw, C.M, Ljung, K., Noel, J.P., and Chory, J.(2014). Генерируемый семядолями ауксин необходим для теневого роста гипокотилей у Brassica rapa . Plant Physiol. 165, 1285–1301. DOI: 10.1104 / стр.114.241844
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рид, Дж. У., Нагпал, П., Пул, Д. С., Фуруя, М., и Чори, Дж. (1993). Мутации в гене фитохрома В рецептора красного / дальнего красного света изменяют удлинение клеток и физиологические реакции на протяжении всего развития Arabidopsis . Растительная клетка 5, 147–157. DOI: 10.2307 / 3869581
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Реймон, М. К., Брюно, Г., Шове, А., Мартинес-Гарсия, Дж. Ф., Мартин-Магнетт, М. Л., Монегер, Ф. и др. (2012). Светорегулируемый генетический модуль был задействован для развития плодолистиков в Arabidopsis после структурного изменения SPATULA. Растительная клетка 24, 2812–2825. DOI: 10.1105 / tpc.112.097915
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Робсон, П., Whitelam, G.C., и Smith, H. (1993). Отдельные компоненты синдрома избегания тени обычно проявляются у мутантов Arabidopsis thaliana и Brassica rapa , дефицитных по фитохрому B. Plant Physiol. 102, 1179–1184.
PubMed Аннотация | Google Scholar
Роиг-Вилланова И., Боу Дж., Сорин К., Девлин П. Ф. и Мартинес-Гарсия Дж. Ф. (2006). Идентификация первичных генов-мишеней передачи сигналов фитохромов. Ранний контроль транскрипции во время реакции избегания тени у Arabidopsis . Plant Physiol. 141, 85–96. DOI: 10.1104 / стр.105.076331
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Роиг-Вилланова, И., Бу-Торрент, Дж., Галстян, А., Карретеро-Паулет, Л., Портолес, С., Родригес-Консепсьон, М., и др. (2007). Взаимодействие избегания тени и реакции на ауксин: роль двух новых атипичных белков bHLH. EMBO J. 26, 4756–4767. DOI: 10.1038 / sj.emboj.7601890
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Роуз, Дж.К., Браам, Дж., Фрай, С. К., и Нишитани, К. (2002). Семейство ферментов XTH, участвующих в эндотрансглюкозилировании и эндогидролизе ксилоглюкана: текущие перспективы и новая унифицированная номенклатура. Physiol растительных клеток. 43, 1421–1435. DOI: 10.1093 / pcp / pcf171
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Солтер, М. Г., Франклин, К. А., и Уителам, Г. К. (2003). Стробирование быстрой реакции избегания тени по циркадным часам у растений. Природа 426, 680–683.DOI: 10.1038 / nature02174
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Sasidharan, R., Chinnappa, C.C., Staal, M., Elzenga, J. T., Yokoyama, R., Nishitani, K., et al. (2010). Опосредованное качеством света удлинение черешка у Arabidopsis во время избегания тени включает модификацию клеточной стенки с помощью ксилоглюкановой эндотрансглюкозилазы / гидролаз. Plant Physiol. 154, 978–990. DOI: 10.1104 / стр.110.162057
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сасидхаран Р., и Пиерик, Р. (2010). Модификация клеточной стенки с участием XTHs контролирует опосредованное фитохромом удлинение черешка у Arabidopsis thaliana . Завод Сигнал. Behav. 5, 1491–1492. DOI: 10.4161 / psb.5.11.13643
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Sessa, G., Carabelli, M., Sassi, M., Ciolfi, A., Possenti, M., Mittempergher, F., et al. (2005). Динамический баланс между активацией и репрессией гена регулирует реакцию избегания тени у Arabidopsis . Genes Dev. 19, 2811–2815. DOI: 10.1101 / gad.364005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Смит, Х. (1982). Качество света, световосприятие и стратегия растений. Annu. Rev. Plant Physiol. 33, 481–518. DOI: 10.1146 / annurev.pp.33.060182.002405
CrossRef Полный текст | Google Scholar
Смит, Х. и Уайтелэм, Г. (1997). Синдром избегания тени: множественные реакции, опосредованные множеством фитохромов. Plant Cell Environ. 20, 840–844. DOI: 10.1046 / j.1365-3040.1997.d01-104.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сорин, К., Салла-Мартрет, М., Бу-Торрент, Дж., Роиг-Вилланова, И., и Мартинес-Гарсия, Дж. Ф. (2009). ATHB4, регулятор избегания тени, модулирует гормональный ответ у проростков Arabidopsis . Plant J. 59, 266–277. DOI: 10.1111 / j.1365-313X.2009.03866.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Тао, Ю., Феррер, Дж. Л., Льюнг, К., Пойер, Ф., Хонг, Ф., Лонг, Дж. А. и др. (2008). Быстрый синтез ауксина по новому триптофан-зависимому пути необходим для предотвращения тени у растений. Cell 133, 164–176. DOI: 10.1016 / j.cell.2008.01.049
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Торнбер, Дж. П. (1975). Хлорофилл-белки: компоненты светоуборочного и реакционного центра растений. Annu. Rev. Plant Physiol. 26, 127–158. DOI: 10.1146 / annurev.стр.26.060175.001015
CrossRef Полный текст | Google Scholar
Варнасоория, С. Н., Брутнелл, Т. П. (2014). Повышение урожайности трав при высокой плотности посадки за счет инженерных световых решений: от модельных систем до исходного сырья. J. Exp. Бот. 65, 2825–2834. DOI: 10.1093 / jxb / eru221
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Яновский, М. Дж., Касал, Дж. Дж., И Уителам, Г. К. (1995). Фитохром A, фитохром B и HY4 участвуют в ответах роста гипокотилей на естественную радиацию у арабидопсиса : слабая деэтиоляция мутанта phyA под плотным покровом. Plant Cell Environ. 18, 788–794. DOI: 10.1111 / j.1365-3040.1995.tb00582.x
CrossRef Полный текст | Google Scholar
% PDF-1.4 % 1 0 объект > эндобдж 9 0 объект /Заголовок /Тема / Автор /Режиссер / Ключевые слова / CreationDate (D: 20211011154752-00’00 ‘) / PDFВерсия (1.6) / ModDate (D: 201152505 + 02’00 ‘) >> эндобдж 2 0 obj > эндобдж 3 0 obj > эндобдж 4 0 obj > эндобдж 5 0 obj > транслировать 2019-06-22T09: 56: 38 + 05: 30Microsoft® Word 20102019-09-30T15: 25: 05 + 02: 002019-09-30T15: 25: 05 + 02: 00application / pdfuuid: 865da925-6c13-4d69-969a -4b0f6c7c19aduuid: ab3c5867-dfbe-4ac3-ab31-2455bc4f3c38www.ilovepdf.com; изменено с помощью iText® 5.4.2 © 2000-2012 1T3XT BVBA (AGPL-версия) 1.6 конечный поток эндобдж 6 0 obj > эндобдж 7 0 объект > эндобдж 8 0 объект > эндобдж 10 0 obj > эндобдж 11 0 объект > эндобдж 12 0 объект > эндобдж 13 0 объект > эндобдж 14 0 объект > эндобдж 15 0 объект > эндобдж 16 0 объект > эндобдж 17 0 объект > эндобдж 18 0 объект > эндобдж 19 0 объект > эндобдж 20 0 объект > эндобдж 21 0 объект > эндобдж 22 0 объект > эндобдж 23 0 объект > эндобдж 24 0 объект > эндобдж 25 0 объект > эндобдж 26 0 объект > эндобдж 27 0 объект > эндобдж 28 0 объект > эндобдж 29 0 объект > эндобдж 30 0 объект > эндобдж 31 0 объект > эндобдж 32 0 объект > эндобдж 33 0 объект > эндобдж 34 0 объект > эндобдж 35 0 объект > эндобдж 36 0 объект > эндобдж 37 0 объект > эндобдж 38 0 объект > эндобдж 39 0 объект > эндобдж 40 0 объект > эндобдж 41 0 объект > эндобдж 42 0 объект > эндобдж 43 0 объект > эндобдж 44 0 объект > эндобдж 45 0 объект > эндобдж 46 0 объект > эндобдж 47 0 объект > / ProcSet [/ PDF / Text / ImageC / ImageB / ImageI] >> эндобдж 48 0 объект > транслировать x ڝ XɎ7 +, Q; C8HnFЋ; Op% VOxS (z |] / Zr / ~ _ / ˇ ~ q 1rǼ ߖ CryMuyqt.